胆蛋白质
胆蛋白质叶绿体色素被发现只有在某些藻类:红藻门、隐藻门和蓝藻门。他们要么是红色(藻红蛋白,phycoerythrocyanin)或蓝色(藻青蛋白,allo-phycocyanins)的颜色。他们已经看过的Bogorad(1975),甘特(1975、1977),O 'Carra和O是什么Eocha(1976),格雷泽(1981、1985),MacColl和Guard-Friar(1987),罗文(1989)和Mimuro和菊池(2003)。
红色和蓝绿藻的胆蛋白质是密切相关的,我们将考虑在一起。他们分为四类的基础上吸收带的位置:这些是为了增加波长,藻红蛋白,phycoerythrocyanins,
200 4 71 im SQO HO) 0
Wavrlrnnrh公里)
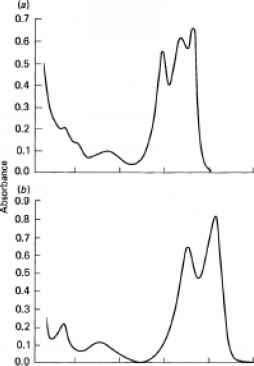
图8.14吸收光谱的胆蛋白质(O是什么Eocha之后,1965)。(一)从烟台R-phycoerythrin石。(b)从Porphyra R-phycocyanin laciniata。
200 4 71 im SQO HO) 0
Wavrlrnnrh公里)
图8.14胆蛋白质的吸收光谱(O是什么Eocha之后,1965)。(一)从烟台R-phycoerythrin石。(b)从Porphyra R-phycocyanin laciniata。
藻青蛋白和allophycocyanins。主要的位置吸收峰表8.4中列出,吸收光谱的红藻藻红蛋白、藻青蛋白图8.14所示。
的一般原则管理这些色素的分布似乎allophycocyanin和藻青蛋白的一个或其它发生在所有红色和蓝绿藻类物种:一个藻红蛋白(或phycoer-ythrocyanin在某些蓝绿藻)可能或不可能,此外,。红藻的藻红蛋白很少缺席,但经常缺席的蓝绿藻。典型的情况是,在红藻类、藻红蛋白构成的大部分胆蛋白质,而在蓝绿色藻类、藻青蛋白或(不常见)藻红蛋白(或phycoerythrocyanin)是主要的组件:allophycocyanin几乎总是一个较小的组件。
藻红素(半胱氨酸残基)
HOOC羧基
HOOC羧基
藻青素(半胱氨酸残基)
图8.15结构和可能的模式结合蛋白,藻胆色素的生色团的藻胆蛋白质。458的689系统的共轭双键,生色团的光谱属性依赖,强调了沉重的线。Phycourobilin可能是CH2而不是CH组加入环B和C, d .连杆通过乙烯基半胱氨酸的蛋白质也可以发生/乙基环D.458
胆蛋白质的生色团欠他们的颜色开四吡咯化合物被称为藻胆色素。有四个主要生色团——藻青素(蓝色),藻红素(红色),phycourobilin(黄色)和phycoviolobilin(紫色)——也称为crypto-violin。不像叶绿素和类胡萝卜素,它们是蛋白质共价结合。他们的链接结构和模式的载脂蛋白在图8.15所示。
所有的藻胆蛋白质包含克分子数相等的数量的两种亚基,称为a和b:一些藻红蛋白包含除了数量小得多(一个分子每6 a或b)第三类的亚基,g。a和b亚基分子量约为17 - 22 kDa但g亚基可能更大。每个a或b亚基胆蛋白质至少有一个藻胆色素发色团。亚基可以有一个或两个发色团;b亚基有一个、两个或三个;和g单元有四个。发色团的分布在两个亚单元在不同的蛋白质表8.5所示。这个分布可能并不是不变的。从红藻藻红蛋白Callithamnion罗森看来phycourobilin随光强的藻红素的比例在增长:1495有可能在一些网站内的蛋白质,可以连接这两个发色团的。
ab胆蛋白质发生聚合的基本结构。隔离,不同大小可溶状态,发现最常见
表8.5的光谱性质不同的胆蛋白质和藻胆色素的分布生色团在他们subunits.457‘459’1002‘1008’
1018、1442、458841842844
表8.5的光谱性质不同的胆蛋白质和藻胆色素的分布生色团在他们subunits.457‘459’1002‘1008’
1018、1442、458841842844
胆蛋白质 |
藻typea |
在光合作用吸收最大值rangeb (nm) |
a-subunitc |
b-subunitc |
g-subunitc |
红色和蓝色 |
|||||
藻类 |
|||||
Allophycocyanin |
R、B |
650,(620) |
1 PCB |
1 PCB |
- - - - - - |
Allophycocyanin-B |
R、B |
671年,618年 |
1 PCB |
1 PCB |
- - - - - - |
C-Phycocyanin |
R、B |
620年 |
1 PCB |
2印刷电路板 |
- - - - - - |
R-Phycocyanin |
R |
618年,553年 |
1 PCB |
1 PCB, |
- - - - - - |
1钢 |
|||||
Phycoerythrocyanin B |
(590),568 |
1的简历 |
2印刷电路板 |
- - - - - - |
|
C-Phycoerythrin |
B |
562 - 565 |
2钢 |
3钢 |
- - - - - - |
CU-Phycoerythrin |
B |
490 - 500,545 - 560 |
2钢 |
3钢, |
- - - - - - |
1酒吧 |
|||||
b-Phycoerythrin |
R |
(563),545 |
2钢 |
4钢 |
- - - - - - |
B-Phycoerythrin |
R |
(565),546,(498) |
2钢 |
3钢 |
2钢, |
2酒吧 |
|||||
R-Phycoerythrin |
R |
568、540、598 |
2钢 |
2钢, |
1钢, |
1酒吧 |
3酒吧 |
||||
隐芽植物 |
|||||
藻青蛋白- 615 |
615年,585年 |
1 PCB |
2印刷电路板, |
- - - - - - |
|
1的简历 |
|||||
藻青蛋白- 630 |
630年,585年 |
留言。 |
留言。 |
- - - - - - |
|
藻青蛋白- 645 |
645,(620),585 |
1 u |
2印刷电路板, |
- - - - - - |
|
1的简历 |
|||||
藻红蛋白- 544 |
(565),544 |
2钢, |
钢 |
- - - - - - |
|
2 CV |
|||||
藻红蛋白- 555 |
555年 |
钢 |
钢 |
- - - - - - |
|
藻红蛋白- 568 |
568年 |
钢 |
钢 |
- - - - - - |
|
一个R,红藻;B,蓝绿藻。b小调最大值或肩膀在括号中。
c PCB,藻青素;钢、藻红素;酒吧,phycourobilin;简历,cryptoviolin;U,未知藻胆色素发色团,有别于前四,峰值在697海里。没有数量意味着生色团的数量是未知的。留言。,没有数据。
似乎六聚体a6 b6和三聚物a3 b3。那些有一个g的藻红蛋白亚基可能发生a6 b6g聚集在孤立的状态。体内,胆蛋白质发生较大的骨料,藻胆体,颗粒直径30到40 nm附加到类囊体的外表面(图8.2)。这些都是有序的,具体的结构
-
.")
- 图8.16藻胆体的示意图表示结构(格雷泽后,1985;甘特,1986;格雷泽和梅丽莎,1987)。
.")
而不是随机骨料,似乎三个左右不同胆蛋白质存在于藻类都是存在于每个藻胆体,加上少量的一些无色蛋白质。Allophycocyanin,稍后我们将看到转移能量收获藻胆体的叶绿素,被认为是位于底部的粒子,相邻细胞膜:另一方面,在Allophycocyanin更充足的蛋白质形成一个外壳,与旁边的藻青蛋白被Allophycocyanin和藻红蛋白或phycoerythrocyanin形成粒子的外围。电子显微镜和生物化学的研究已经引起phyco-bilisome模型结构的示意图见图8.16。在叶绿素d-containing原核生物,Acaryochloris码头,胆蛋白质(藻青蛋白和allophycocyanin)作为杆状结构存在类囊体membrane.594位于细胞质的一面
转向隐藻门我们找到一些相似之处,以及显著的区别,红藻门和Cyano-phyta的情况。cryptophytan色素红色或蓝色,相应地分为藻红蛋白和藻青蛋白。每个隐芽植物物种似乎只包含一个胆蛋白质。没有allophycocyanin-like颜料尚未被发现在这一组。主要吸收峰是在612到645纳米范围内的藻青蛋白,和544到568纳米范围的藻红蛋白。到目前为止的生色团中发现这些蛋白藻红素,藻青素,cryptoviolin (phycoviolobilin)和一个未知的藻胆色素的吸收峰在697 nm.580(酸性尿素)
组织的隐芽植物胆蛋白质似乎在同一尾红和蓝绿藻类蛋白质亚基原则:在孤立的国家它们的存在主要是a2b2二聚体。子单元,分别为10和17.5 kDa,出现,然而,比在其他藻类更小,并且有两种类型的亚基,使二聚体组合是更好地表示为aa 'b2.513从哪里不同最引人注目的是相应的蛋白质在其他藻类是光合器的位置:代替目前的藻胆体表面的类囊体,该隐芽植物胆蛋白质发生在一个密集的细粒度矩阵填充的内部类囊体(图8.3)。
继续阅读:反应中心和能量转移
这篇文章有用吗?