干旱地区的植物多样性
能够承受长时间的水分短缺并不是任何一种植物群或生命形式的特权。在乔木、灌木、草、蕨类、苔藓和地衣中,有一些物种是干旱耐受性(图2.21-2.22)和其他局限于水域丰富的植物。在所有主要的植物生命形式中,这种抗旱物种的进化有时会被忽视,因为人们很容易被从高海拔地区到低海拔地区植被发生的巨大变化所欺骗低降雨量.特别是,树木覆盖率的减少,在到达干旱地区时可能会给人留下非常生动的印象,这可能会导致低估某些树种的耐旱性范围。
在许多干旱地区,树木的缺乏既是由于降水的减少,也是由于人和动物的原因。当进入雨影面积从降水充足之一,减少水的供应对森林物种组成的影响大于对树木形态存在的影响。原产于干旱地区的树种不一定比草原地区消耗更多的水。在智利和巴塔哥尼亚西部凉爽的雨林里降雨的支持常绿南方山毛榉(Nothofagus dombeyii)的生长需要2500-3800毫米。
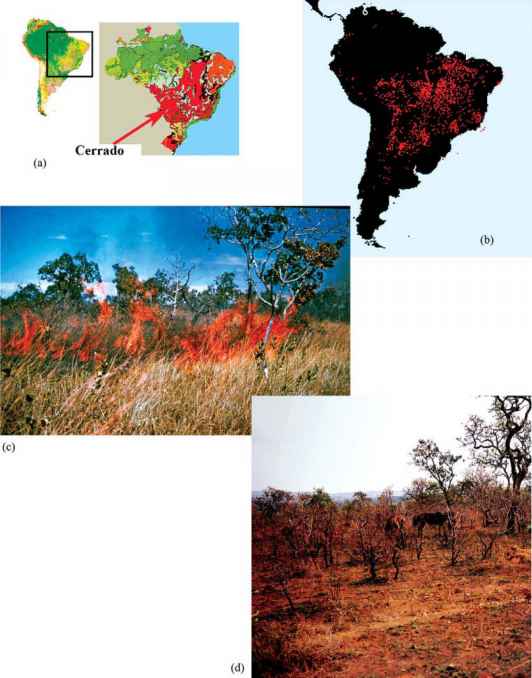
图2.20火与塞拉多。塞拉多的地理位置。(b)根据卫星探测夜间热点记录的2003年9月南美洲火灾发生率(资料来源:世界火灾地图集-欧洲空间局网页:http://dup.eserin.esa.int /爱奥尼亚/。(c)塞拉多植被燃烧(P. E. Gibbs博士摄)。(d)火灾后塞拉多-注意最近火灾产生的扭曲变黑的茎,从中将出现新的嫩枝。
图2.20火与塞拉多。塞拉多的地理位置。(b)根据卫星探测夜间热点记录的2003年9月南美洲火灾发生率(资料来源:世界火灾地图集-欧洲空间局网页:http://dup.eserin.esa.int /爱奥尼亚/。(c)塞拉多植被燃烧(P. E. Gibbs博士摄)。(d)火灾后塞拉多-注意最近火灾产生的扭曲变黑的茎,从中将出现新的嫩枝。

图2.21在巴林(波斯湾)沙漠中的豆科植物(豆科植物)古代标本。这个非凡的标本在当地被称为生命之树。该物种原产于中美洲和美国南部各州,但现在广泛分布在其他干旱地区。在一些地区,如在美国,它被认为是一种入侵灌木,由于过度放牧和气温升高,它正在接管以前的草地。在其他地方,如波斯湾,它的价值在于它的沙粒结合性能,遮荫,可食用的水果,以及作为蜂蜜的来源。
图2.21在巴林(波斯湾)沙漠中的豆科植物(豆科植物)古代标本。这个非凡的标本在当地被称为生命之树。该物种原产于中美洲和美国南部各州,但现在广泛分布在其他干旱地区。在一些地区,如在美国,它被认为是一种入侵灌木,由于过度放牧和气温升高,它正在接管以前的草地。在其他地方,如波斯湾,它的价值在于它的沙粒结合性能,遮荫,可食用的水果,以及作为蜂蜜的来源。
往东60公里雨影安第斯山脉的山脉使降雨量减少到不足350毫米,然而尽管如此,树木仍然在没有放牧的地区存活。
的肉质植物是干旱地区最具特色的植物类群之一。尽管它们被限制在有限的家族中,并且在采用进行石笋酸代谢(CAM)的能力方面具有同质性,但它们的形态却惊人地不同,生长形式可以是树木也可以是灌木。众所周知的约书亚树(Yucca brevifolia)是显示CAM代谢的较大物种之一(图2.22)。只要有一些水分,即使只是沿海的雾,几乎总是有一种植物适应利用生态窗口,无论多么短暂。炎热的沙漠通常是古老的栖息地,生活在那里的植物和动物有着漫长的干旱适应历史。因此,低纬度干旱地区的生物多样性不足为奇。

图2.22易干旱生境中树木形态的变化。(上)的科迪勒拉山系产于巴塔哥尼亚沙漠(阿根廷)西部边缘的一种柏树(澳洲柏树)。(上图左)莫哈韦沙漠中的乔舒亚树(短叶丝兰)——百合科的一种树状植物,是具有石蜡酸代谢(CAM)的较大物种之一。(上图右)巴拉圭的Ceiba chodatii(邦巴科),一种储存水的瓶子树(摄影:P. E. Gibbs博士)
图2.22易干旱生境中树木形态的变化。(上)巴塔哥尼亚沙漠(阿根廷)西部边缘的科迪勒拉柏树(Austrocedrus chilensis)。(上图左)莫哈韦沙漠中的乔舒亚树(短叶丝兰)——百合科的一种树状植物,是具有石蜡酸代谢(CAM)的较大物种之一。(上图右)巴拉圭的Ceiba chodatii(邦巴科),一种储存水的瓶子树(摄影:P. E. Gibbs博士)
虽然沙漠在气候上可以归为干旱地区,但干旱期的周期性和持续时间有很大差异。任何一个专业的考试沙漠地区揭示了相当大的气候变化。位于智利和秘鲁的阿塔卡马沙漠是世界上最干燥的沙漠之一。在安托法加斯塔,有时在20年里不到6年就能探测到可测量的降雨,然后每年降水量不超过4-6毫米。然而,从5月到11月,一种稠密的冷雾,Garua(秘鲁)或Camancha(智利),伴随着寒冷的海风滚滚而来洪堡当前.它在夜间特别稠密,但在白天足够厚,可以一次遮蔽几天的热带太阳。在8月份,一些暴露在这种雾中的地区只有36小时的日照,平均温度为13°C,而在雾上方不到800米的上坡处,温度急剧上升到24°C。这种薄雾支撑着以地衣为主的植被,覆盖着面向大海的大雾笼罩的大面积山丘。
在与阿塔卡马沙漠接壤的一些特别受欢迎的地方,有一个独特的森林植被,洛玛斯。树下的降水量是户外凝结的8倍。典型树种有Carica candens,各种Eugenia, Caesalpina tinctoria和Schinus molle,以及靠近Lachay的Acacia macrocantha (Hueck, 1966)。洛玛有很高比例的特有物种和丰富的凤梨植物区系。
在阿塔卡马沙漠中同样引人注目的是14种Tillandsia,既有附生的,也有无根的,陆地代表(Rundel & Dillon, 1998)。Rundel和Dillon列出的所有Tillandsia物种都是广义上的附生植物,但除了生长在植物上外,它们也生长在岩石上,其中6个物种(T. purpurea, T. latifolia, T. capillaris, T. marconae, T. werdermanii和T. landbeckii)都进化出了在沙子上不生根生存的能力(图2.23)。正如朗德尔所指出的那样,世界上没有任何地方的凤梨比这些生长在沙滩上的沿海物种更具优势或具有更多的生物量。这些物种中的许多生长在维管植物耐受的绝对极限。有可能找到一个由单一Tillandsia物种组成的整个群落。
撒哈拉沙漠位于非洲,面积近900万平方公里,年平均降雨量不足100毫米。然而,尽管总体干旱,但生物气候的多样性相当大。在北部的地中海撒哈拉沙漠和南部的热带撒哈拉沙漠之间,还有中原撒哈拉沙漠、山地撒哈拉沙漠和大洋撒哈拉沙漠,它们都有自己独特的气候模式和生物多样性。分布模式动植物的变化与气候参数密切相关,特别是在降雨量和温度的数量和季节性方面。后者在控制动植物分布方面可能与前者发挥同样重要的作用,因为在热带降雨制度下,地中海物种可以在较高海拔的群落中占主导地位,而热带物种则侵入冬季温度足够温暖的地中海降雨地区(Lehouerou, 1995)。
非洲的沙漠也提供一些令人惊讶的例子,证明只要降水有规律地发生,维持一个物种丰富的植物群所需的降雨量是多么少。南非纳马夸兰西北海岸诺洛思港和亚历山大湾之间的沙质海岸带是生物多样性多肉卡鲁生物群落的一部分。在750平方公里的区域内,有300种植物,24%的地方率(Desmet & Cowling, 1999)。
埃及西奈半岛有大约1285种植物。南部地区物种尤其丰富,有800种(包括种下类群),特有种约占4.3%。圣凯瑟琳地区不同地貌之间的贝塔多样性一方面反映了斜坡和阶地之间以及阶地和山脊之间的巨大生物变化(Ayyad et al., 2000)。到目前为止,分子遗传学研究在沙漠很少,但已经进行的研究表明种群之间存在相当大的多样性。在某些情况下,这可能与长寿许多沙漠多年生植物。对圣凯瑟琳沙漠中昆虫授粉的沙漠物种东方Alkanna orientalis的遗传亚结构的调查显示,生长在三个不同陡峭的溪谷和一个中央平原地区的亚种群在遗传上彼此不同,但仍然显示出基因流动的证据,特别是在两个溪谷和邻近的平原之间(Wolff et al., 1997)。圣凯瑟琳沙漠的河谷之间的高山山脊使得蜜蜂不太可能在山脊上移动,蜜蜂也不可能在每条河谷的几株植物范围之外的地方觅食。因此,山洪传播的种子似乎更多
-
 in the Atacama Desert. This is a free-living bromeliad species that survives unrooted and moves about the desert blown by the wind.")
- 图2.23阿塔卡马沙漠中的凤梨科Tillandsia latifolia。这是一种自由生活的凤梨科植物,没有根,随风在沙漠中移动。
 in the Atacama Desert. This is a free-living bromeliad species that survives unrooted and moves about the desert blown by the wind.")
这可能是种群间基因流动的原因,并再次说明,即使在环境紧张的栖息地,也可以从干扰中获得一些好处。尽管炎热的沙漠在空间上和时间上明显空虚,但其中却包含着物种丰富多样的植物区系,这再次说明边缘地区也可以拥有显著的生物多样性。
继续阅读:非生产性栖息地的资源必需品
这篇文章有用吗?


