早期后生动物的同种异体的工程师
没有垂直生物扰动作用在硅质碎屑的岩石沉积在新元古代和早寒武纪表明条件下海底表面可能是不利的后生动物的活动(Bottjer et al . 2000年)。尽管沉积物内有限的食物资源提供了激励生物洞穴infaunally,相当多的证据显示,其中包括丰富的微生物介导的沉积结构的存在,表明身体和适应性的局限性主要是负责限制底栖生物epifaunal栖息地(例如,Bottjer et al . 2000年)。
生物扰动作用的化石记录展品突出的趋势随着时间的推移对痕迹化石的复杂性增加,密度,穿透深度下的海底(Ausich Bottjer 1982 Droser和Bottjer 1993)。最早期的宏观痕迹化石被发现在岩石沉积在新元古代末期(例如,詹森2003年,詹森et al . 2006年)。大多数这些早期的生物结构由简单的双边对称的,水平的活动形式,可能代表软体蠕虫状的生物在海底表面或下微生物垫(柯林斯等。2000年,1995年情人节)。跟踪化石才开始表现出一个垂直面向组件Neoproterozoic-Cambrian边界(ca 5.42亿年前),当Treptichnus pedum,由一系列的痕迹化石浅scoop-like标志,出现在岩石代表浅海洋环境(例如,Droser et al . 1999年,格尔et al . 2001年)(图8.3)。尽管深度垂直洞穴发生在近岸和临滨环境在最早的寒武纪(“针管迹piperock”;Droser 1991),浅洞穴结构与很少或没有垂直度是在潮下的环境下生物扰动作用的主要形式,直到中间寒武纪末(Bottjer et al . 2000年)(图8.3)。生物扰动作用的逐渐增加深度浅从早期到晚期寒武纪海洋环境已经证明了Droser(1987)和Droser Bottjer (1988、1989)。沉积的岩石浅海洋设置寒武纪通过现代的展示一个非常不同的从这些代表寒武纪早期海床特征,包括明显中断沉积地层,常见的垂直洞穴可能套印生物扰动作用的结构,和缺席microbi-ally介导沉积结构(Bottjer et al . 2000年)(图8.3)。因此,转变发生在海底之间的寒武纪主要表现为水平生物扰动作用和广泛的微生物垫(反映在丰富的微生物介导的沉积结构和缺乏垂直沉积中断低寒武纪岩石)和那些被广泛的垂直特征生物扰动作用和微生物垫缺席。
Seilacher和弗鲁格(1994)提出了农业革命假说来解释如何以及为什么这种转变在海底条件和底栖生物的行为发生。根据这一假说,底栖生物

图8.3痕迹化石保存在较低的寒武纪(A, B)和始新世(ca 56-34百万岁)(C)的岩石。(A) Treptichnus pedum,第一个跟踪化石展览面向垂直的分量,保存颠倒在寒武纪沉积岩单位较低的底部。嵌套叶(箭头)的跟踪可能代表系统探测海底沉积物的priapulid-like deposit-feeding有机体。(B)寒武纪层面较低表面含有丰富的水平跟踪化石(Planolites;箭头)。(C)丰富的垂直导向的痕迹化石在圣地亚哥附近的始新世曝光,CA。Conostichus(黑色箭头),一个大型的、lobe-shaped洞穴,是由海葵和其他固定底栖生物悬浮泥沙涌入期间喂食器。Ophiomorpha(白色箭头),深mud-lined洞穴,是由许多类型的底栖生物suspension-feeding甲壳类动物,需要稳定的非永久性的住所。(见色板)。
图8.3痕迹化石保存在较低的寒武纪(A, B)和始新世(ca 56-34百万岁)(C)的岩石。(A) Treptichnus pedum,第一个跟踪化石展览面向垂直的分量,保存颠倒在寒武纪沉积岩单位较低的底部。嵌套叶(箭头)的跟踪可能代表系统探测海底沉积物的priapulid-like deposit-feeding有机体。(B)寒武纪层面较低表面含有丰富的水平跟踪化石(Planolites;箭头)。(C)丰富的垂直导向的痕迹化石在圣地亚哥附近的始新世曝光,CA。Conostichus(黑色箭头),一个大型的、lobe-shaped洞穴,是由海葵和其他固定底栖生物悬浮泥沙涌入期间喂食器。Ophiomorpha(白色箭头),深mud-lined洞穴,是由许多类型的底栖生物suspension-feeding甲壳类动物,需要稳定的非永久性的住所。(见色板)。
后生动物的进化适应的寒武纪大爆发,获得允许他们向matgrounds垂直洞穴。Bio-turbation深度和强度增加,最终破坏微生物垫的分层结构,增加水和氧含量的海底沉积物。垫开发被降级边缘环境的农业革命,和海底post-Cambrian海洋环境特征更典型,如改善营养分布和模糊watersediment边界(Bottjer et al . 2000年)(图8.1)。
生态和进化的农业革命的影响反映在身体和痕迹化石的记录和集体被称为寒武纪衬底革命(Bottjer et al . 2000年)。更重要的这些影响是那些感到垫底社区(Seilacher 1999)。垫刮子和undermat矿工更好比其他公会为适应新的海底条件,因为他们的移动生活方式变化而让他们重新定位自己在氧气和衬底一致性(Bottjer et al . 2000年)。然而,垫刮子适应生活和喂养凝聚力沉积物表面,和失踪的表面从开放海洋环境迫使许多物种迁移到更硬基质限制地区是常见的,如多岩石的海岸线和深海(Bottjer et al . 2000年)。垫encrusters和垫贴纸面临更大的挑战由于他们专门的固着生活方式。缺乏一种迁移到更合适的环境中,许多这样的团体发展茎或直接附件机制,允许他们利用有限的硬表面,可用在浅海洋环境(Bottjer et al . 2000年)。然而,并不是所有的这些组织是成功的。的mat-sticking helicoplacoid棘皮动物,例如,不适应新的基质条件和灭绝在年底前寒武纪(Bottjer et al . 2000;Dornbos Bottjer 2000, 2001)。
农艺和寒武纪基质最早期革命一起代表同种异体的实例生态系统工程后生动物的生命的历史。随着生物扰动作用的深度和强度增加,底栖生物后生动物带来了戏剧性的变化在寒武纪早期浅潮下的海床,取代微生物作为主要生物影响许多海底条件和提供社区的其他成员各种前所未有的资源和生态位(Bottjer et al . 2000年,Dornbos et al . 2004年)(图8.4)。bioturba-tion已经承认变革作用的在各种各样的现代生态系统(例如,Meysman et al . 2006年)。
在这种情况下外源的生态系统工程,如前面讨论的一些例子,工程师自己都不一定保留,但工程活动的影响可以很容易地认出了岩石中。努力确定生态系统工程师(s)的农业革命是在早期阶段,虽然软体后生动物可能是基于他们的丰度异常保留存款如澄江生物群(侯et al . 2004年)。鉴于保存软组织的稀缺的化石记录,研究痕迹化石的分布和丰富低寒武纪岩石物理物理
Post-Cambrian
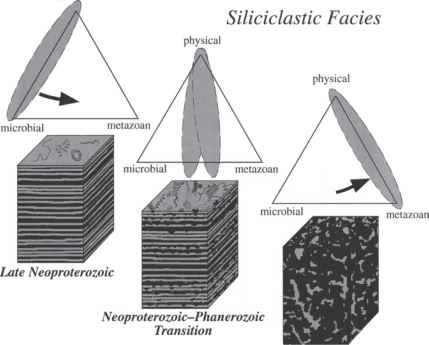
图8.4的示意图说明转变Neopro-terozoic末和post-Cambrian之间发生的主要过程,海底条件控制。表示在三角图,晚新元古代海底条件由物理和微生物过程控制。微生物的影响逐渐减少Neoproterozoic-Phanerozoic过渡期间,当后生动物bioturba-tion变得更加丰富和破坏性的。post-Cambrian,后生动物的丰富和广泛的生物扰动作用的主要因素,除了物理过程,海底控制条件。修改Bottjer et al。(2000)。
Post-Cambrian
图8.4的示意图说明转变Neopro-terozoic末和post-Cambrian之间发生的主要过程,海底条件控制。表示在三角图,晚新元古代海底条件由物理和微生物过程控制。微生物的影响逐渐减少Neoproterozoic-Phanerozoic过渡期间,当后生动物bioturba-tion变得更加丰富和破坏性的。post-Cambrian,后生动物的丰富和广泛的生物扰动作用的主要因素,除了物理过程,海底控制条件。修改Bottjer et al。(2000)。
可能是最好的方法来确定角色的软体生物可能在寒武纪早期工程生态系统。研究低寒武纪浅海洋岩石在加州东部表明简单的水平跟踪化石Planolites可能浅钻洞的产品软体蠕虫状的后生动物,是最丰富的一种生物扰动作用在水平“层面”在岩石表面检查(Marenco和Bottjer出版社)。Planolites扩散在寒武纪早期海底洞穴可能反映了存在稳定的营养供应,所产生的广泛的微生物垫,这可能是维持一个多样化matground社区的能力。其他证据存在的这样一个社区在这些低寒武纪岩石单元包括Volborthella常见的一个小神秘寒武纪化石解释为骨架的matground-adapted动物(例如,Seilacher 1999)和壳牌石膏模型、模具可能linguliform腕足类,可能已经适应了生活在低氧条件下被微生物垫(贝利et al . 2006年)。尽管丰富的水平生物扰动作用,微生物活动很可能仍然在这些特定的主导因素影响基质条件早前寒武纪浅海洋环境的农业革命(贝利et al . 2006年)。
继续阅读:Sphaeroma Quoianum
这篇文章有用吗?