叶绿体运动
光的速度收集叶绿体不仅取决于他们的颜料补还细胞内的位置和姿态。在某些水生植物随光强度。在高plants1415行为的一般模式,在低强度光等细胞叶绿体移动一个位置光吸收是最大化——他们传播自己的,平行,那些面对的细胞壁入射光。在高强度光(饱和光合作用),叶绿体搬到一个位置,这样光吸收减少,可能减少光抑制:他们远离细胞壁直接暴露在光和结盟的,相邻平行,那些最阴影的细胞壁,典型的侧墙,这样它们不仅侧面而不是正面光,但也有些阴影。水生的被子植物Potamogeton管,在苏格兰尼斯树叶收集从0.25深度的叶绿体在侧墙的细胞,而叶子从2.5面临的表面深度的分布在叶绿体光。1273的另一种方式减少光在高强度是叶绿体原子核周围组装在一个集群中:这是观察叶子的海草Halophila stipulacea增长0.5米深度Mediterranean.323沙龙和啤酒(2008)相比,h . stipulacea植物生长在高光(450更易与光子m ~ 2中午s_1)或低光(150 m ~ 2中午s_1)更易与光子。在日出前h 06:00时,叶绿体是均匀分散的叶细胞的细胞质内。高亮的植物,叶绿体的迁移从日出开始,开始观察凝结在喂饲h。完全凝结发生了中午和持续整个下午。在下午晚些时候叶绿体开始分散在细胞质中,实现和充分分散在日落之后几小时。在低光植物叶绿体白天没有凝结。高亮的植物叶绿体凝结伴随着减少叶子吸收比清晨从0.56到0.34,中午没有任何附带的叶绿素含量的变化。在植物的h . stipulacea阿卡巴湾(红海),施瓦兹和Hellblom(2002)发现,植物叶绿体凝结在7米深度叶吸收率下降到55%,85%的植物在30米。
的光谱在高等植物叶绿体运动有一个峰值在蓝色区域约450海里:感光可能黄素或类胡萝卜素。544年的机制可能与细胞质流:叶绿体似乎和细胞质而不是通过一起移动。
Nultsch和Pfau(1979)研究了大量的叶绿体运动沿海和近岸的海洋macroalgae。布朗在大多数的藻类物种,叶绿体搬到面临的细胞壁低照度的光和侧墙平行光的方向在高辐照度(图12.19):从一个位置到另一个1到2 h,没有明确的光致运动观察叶绿体的绿色或红色藻类物种进行了研究。在siphonac-eous绿藻Caulerpa racemosa,生长在热带浅水礁地区,Horstmann(1983)观察到,在明亮的阳光下chloro-plasts收回从叶子到匍匐枝。
-
, Marine Biology, 51, 77-82). Arrangement of the chloroplasts in (a) low-intensity (1000 lux) and (b) high-intensity (10 000 lux) light. Magnification ~4000 x.")
- 图12.19光致运动叶绿体的褐藻植物saccharina (Nultsch许可和Pfau(1979),海洋生物学,51岁,77 - 82)。叶绿体的安排(a)低强度(1000勒克斯)和(b)高强度(10 000勒克斯)光。放大~ 4000 x。
, Marine Biology, 51, 77-82). Arrangement of the chloroplasts in (a) low-intensity (1000 lux) and (b) high-intensity (10 000 lux) light. Magnification ~4000 x.")
丝状绿藻转板藻和Mesotaenium,单一,平集中位于矩形在每个细胞叶绿体,叶绿体的特征运动响应的变化是将光政权纵轴,而不是移动墙上。结果正面温和的光,和侧面强烈的光。Haupt(1973)和同事表明感光控制这个运动是光敏色素。在另一个丝状,合胞体的海藻,Vaucheria sessilis (Xanthophyceae),如果灯丝发光一度与低强度蓝光,叶绿体等细胞器,通常携带流细胞质造成细胞的总照明部分。125的photosyn-thetic影响还不清楚。
浮游藻类在浮动的情况下,随机取向的光,运动本身的一部分细胞或其他细胞不可能——如果只包含一个或几个叶绿体产生多大的影响,光的速度收集。在某些情况下,chloro-plast可以减少吸收截面在明亮的光线萎缩:这是观察dinoflagellate1329和硅藻。537686年海洋中心硅藻Lauderia北欧化工,大约有50个每细胞叶绿体,基弗(1973)观察到的前2分钟暴露在强烈的光(244 Wm2)叶绿体简约的大小(图12.20)。叶绿体在接下来的30到60分钟,在低亮度均匀分布在外围,搬到valvar目的细胞的形成两个等大的聚集(图12.20 b)。叶绿体的大小和位置的变化,伴随着减少约40%的吸光度的悬挂在440 nm(可能低估了由于散射总是贡献一些虚假的吸光度):因此,收缩的组合和聚合这种硅藻可以大大减少其能源收集在明亮的光线。
史蒂芬斯(1995)研究了光致叶绿体迁移,和相关的光谱变化,腰鞭毛虫,Pyrocystis甲半月,一个物种丰富的热带和亚热带地区的海洋,拥有大型新月形的细胞。细胞生长在12 h: 12 h暗周期。在暗循环叶绿体是位于胞质链中细胞的两个远端部分(新月的“角”),和缺席的颗粒细胞的中心区域。当细胞被黑暗和放置在显微镜的光束,叶绿体开始移动和2到3分钟后聚集在中央区域的细胞。的吸收光谱的
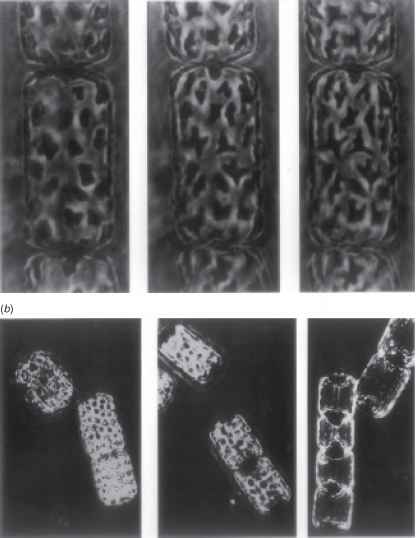
图12.20叶绿体高引起的收缩和聚合光强度在海洋硅藻Lauderia北欧化工(许可,从基弗(1973),海洋生物学,23日39-46)。在每种情况下一系列的显微图显示增加光照改变从左到右。(一)收缩的叶绿体。(b)的运动细胞的叶绿体valvar目的,其次是聚合。
图12.20叶绿体高引起的收缩和聚合光强度在海洋硅藻Lauderia北欧化工(许可,从基弗(1973),海洋生物学,23日39-46)。在每种情况下一系列的显微图显示增加光照改变从左到右。(一)收缩的叶绿体。(b)的运动细胞的叶绿体valvar目的,其次是聚合。

120 144
图12.21光合能力(Pm)的昼夜节律phyto-plankton圣劳伦斯河口,加拿大,在七天时间内(从Demers许可,和勒让德(1981),海洋生物学,64年,243 - 50)。
120 144
图12.21光合能力(Pm)的昼夜节律phyto-plankton圣劳伦斯河口,加拿大,在七天时间内(从Demers许可,和勒让德(1981),海洋生物学,64年,243 - 50)。
聚合叶绿体的细胞表现出更为明显的中心方案的效果,即压扁的山峰,比细胞的叶绿体远地区。在这种情况下,叶绿体运动的影响减少能源的速度收集在明亮的光线。
继续阅读:的microphytobenthos
这篇文章有用吗?
读者的问题
-
挚友1个月前
- 回复
-
布鲁诺1个月前
- 回复
-
伊恩•1个月前
- 回复
-
feaven6个月前
- 回复
-
玛丽亚6个月前
- 回复