珊瑚礁生态系统的变化,气候和全球变化的一个指标raybet雷竞技最新
马丁j . Attrill
海洋研究所海洋生物学和生态学研究中心,德雷克马戏团,普利茅斯大学普利茅斯PL4 8 aa,联合王国
1。介绍4。结论
2。热带珊瑚礁生态系统的引用
3所示。相关的动物群的珊瑚礁
1。介绍
相比与陆地系统和其他组件的海洋环境,相对很少有人知道气候和全球变化是如何影响海底生物的但有一个例外:热带珊瑚礁。raybet雷竞技最新最近的评论[1,2]和metanalyses(3、4)齐心协力所有现有知识如何影响气候,例如,范围变化和物候学[4]的物种,但是,除了珊瑚,一些例子在这些概述来自海洋海底。raybet雷竞技最新帕尔玛在她优秀的回顾,[1]一节致力于海洋社区转变,但大部分例子都来自远洋(第12章)或潮间带区域(第15章);只有两个研究鱼(5、6)相关的潮下的海底。陆地和淡水的例子主导这些研究。相当大比例的研究评估气候对海洋的影响系统的响应主要调查raybet雷竞技最新raybet雷竞技最新气候循环,如厄尔尼诺南方涛动(ENSO)和北大西洋涛动(NAO)。这些提供信息系统如何应对冷却和变暖趋势在这些周期的极端,从而提供一个潜在的生物模型和系统[7]可以应对气候变暖,尤其是当ENSO事件的发生和严重程度预测raybet雷竞技最新
raybet雷竞技最新:观察到的气候变化对地球的影响
版权©2009爱思唯尔帐面价值保留所有权利的任何形式的复制。
气候变暖情景下增加[8]。未来大气动力学和温度变化之间的关系,然而,不确定的,所以还需要小心一些关于过去的生物系统的响应这些气候周期反映了正在进行的和未来的气候变化[1]。raybet雷竞技最新然而,这样的研究提供的信息对气候海洋系统的响应。raybet雷竞技最新
相比其他潮下的底栖生物系统,全球气候变暖的问题对珊瑚礁的影响有一个最高的配置文件近年来,特别是1997年的全球影响1998年极端厄尔尼诺事件[9],公众广泛关注这个生态系统和最近的报告预测普遍亏损的礁石和灭绝的物种(10、11)。raybet雷竞技最新气候变化可能影响珊瑚礁通过几个关键机制,特别是增加海表面温度(SST),海洋酸化,增加猛烈和海平面上升(12、13)。后者三个机制在其他章节专门处理,因此本章将集中在海洋温度的上升对珊瑚礁生态系统的影响,特别是大规模珊瑚本身的漂白效果。
2。热带珊瑚礁生态系统
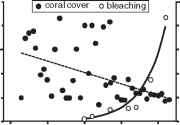
关注人类对珊瑚礁的影响已经存在了几十年,直到最近比较,主要威胁的完整性珊瑚礁生态系一直被认为是过度捕捞和污染(12、14)。这些影响可能在地方一级管理,但任何此类管理时将不成功投入的背景下,最近承认的影响全球气候变化[12]。raybet雷竞技最新同样,可能加剧的气候影raybet雷竞技最新响的附加这些当地其他人为因素的影响,使珊瑚礁系统在世界的一些地区更容易受到伤害。气候变化之间的联系和region-raybet雷竞技最新scale大规模珊瑚白化的现在是无可争议的(12、13),尤其是之间的直接联系漂白和海温异常[15]。没有质量记录漂白在1980年代之前,尽管目前尚不清楚如何广泛等漂白之前是二十世纪初的媒体广泛报道[16];不过不太可能,近年来漂白的规模应该引起注意。在大堡礁,例如,漂白事件已成为1980年代以来更广泛(图1),与珊瑚覆盖下降这一次[17];在全球范围内大规模珊瑚白化变得更加频繁和剧烈的近几十年来[11]。
漂白是由于美白后的珊瑚共生黄藻的驱逐,藻类提供珊瑚大部分的色素。黄藻的损失,因此,叶片相对无色珊瑚组织加上白色的石灰珊瑚礁骨架。这个过程通常被认为是作为应对环境温度高于阈值的增加,^ 0.8 1°C以上夏季平均气温至少4周(18、19)。
60 50 40 30 20 10
60 50 40 30 20 10
0党卫军
1960 1970 1980 1990 2000年
1960 1970 1980 1990 2000年
1008060 -
珊瑚覆盖图1,趋势和珊瑚礁的数量和质量在大堡礁漂白,澳大利亚(适应由麦克米伦出版社有限公司(自然),许可Ref。[17])。b。微分的九种珊瑚漂白反应好,法属波利尼西亚,在2002年5月(重绘Ref。[12],从AAAS允许转载)。疣(anthocercis;b, a婆;c, Montipora tuberculosa;d, Pocillopora疣状;e m . caliculata;f, Leptastrea transversa;g、p . eydouxi;h p . meandrina; i, L. bewickensis; j, Porites lobata; k, L. purpurea).
漂白阈值在珊瑚物种很可能代表了广泛的反应(图1 b),然而,脆弱的感情会随着时间改变后的表型和遗传反应珊瑚[12]。很明显,许多珊瑚物种存在的多种生物地理学的温度和个人随后不同漂白的绝对温度阈值,表明自适应能力在物种[20]。漂白的主要驱动力,因此,似乎是温度上升高于一般经历了珊瑚在任何给定的位置。提出,漂白反应是一个自适应过程(21、22),珊瑚驱逐易感共生体,占用更多的抗药的;虽然有证据表明[13],它似乎并不支持对漂白珊瑚[12]的命运,或许更准确地描述为一种应激反应。
什么是无争用的,然而,是重大漂白事件可能会严重影响珊瑚礁从长远来看:如果漂白是长期或超过2°C以上季节性maxima珊瑚死亡[13]。主要漂白事件中观察到1982 1983,1987 1988,1994 1995,1997 1998 [13],2002
(GBR[23])和2005年(加勒比,[19]),通常,但并非总是如此,伴随着强烈的厄尔尼诺事件,加强全球海洋温度。1997年1998事件是历史上最极端的厄尔尼诺现象[9],并导致广泛的漂白记录世界各地的珊瑚礁[24]。估计世界上16%的珊瑚礁是迷失在这一事件,特别是在印度洋/东南亚[24](表1),只有部分复苏明显。总的来说,只有大约一半的珊瑚礁的影响在1998年恢复[24]。发生2005事件,然而,没有一个厄尔尼诺现象的影响,提供了证据底层增加的趋势在全球海水温度(第十九章);这是有关人为强迫自1970年代以来在大西洋[19]。反常地温暖的气温记录在加勒比海和大西洋热带[19],导致异常水平的漂白:90%的珊瑚覆盖在英属维尔京群岛,美属维尔京群岛的80%和66%在特立尼达和多巴哥,例如[19]。分析当地的温度异常显示,太平洋高于预期年度最大值的时间比先前记录[19](图2),导致观察到的异常漂白。同样值得注意的是,这些极大值已经超过了自1995年以来每年(图2);在此之前,这样的极端罕见。 The second highest value occurred in 1998, when extensive bleaching was also apparent in the Caribbean [25].
唯一可用的长期数据的影响气候对珊瑚物种的地质记录,审查的休斯等。[12]。raybet雷竞技最新许多现存物种的珊瑚可以追溯到时间的上新世(1.8 - 5.3 Ma前),所以经历了广泛和快速的时期在更新世和全新世气候变暖和变冷之前人类的影响[14]。为了应对气候变化,有证据表明物种进行了大的变化在他们的分布范围[26];对珊瑚来说,这个扩展到500公里在澳大利亚南部,例如[12]。直到最近,没有证据表明这些变化以应对现代气候变化,但是,随着海洋温度的增加,两种疣扩大其范围向北50公里沿佛罗里达半岛地区他们没有记录了6000年[27]。raybet雷竞技最新
多的证据,因此,存在增加太平洋通过广泛影响珊瑚礁白化和随后的死亡率,尤其是在特殊的年平均最高气温超过。珊瑚也会影响通过海水pH值的变化,影响他们的能力产生骨骼钙质礁在21章覆盖(见参考。[11]),并有可能进一步影响严重的风暴[28]和[29]海平面上升,气候变化的其他后果[13]。raybet雷竞技最新与过去的气候变化,但是,raybet雷竞技最新珊瑚礁也显著协同效应的影响其他人为活动,如钓鱼和污染,使他们更容易受到变化与当前气候变暖[12]。
/ \表1总结状态的珊瑚礁在17个地区的世界
2004年从Ref。[24],显示比例的珊瑚礁被摧毁(即每个地区。,90%的珊瑚失去和不太可能恢复),加上
地区 |
珊瑚礁区域/平方公里 |
珊瑚礁破坏/ (%) |
珊瑚礁毁于1998 / (%) |
珊瑚礁恢复/ (%) |
||||
这些深渊 |
3800年 |
65年 |
15 |
2 |
||||
南亚 |
19210年 |
45 |
65年 |
13 |
||||
东南亚 |
91 700 |
38 |
18 |
8 |
||||
西南印度洋 |
5270年 |
22 |
41 |
20. |
||||
加勒比 |
3040年 |
16 |
NA |
美国 |
5120年 |
15 |
NA |
NA |
E & N亚洲 |
5400年 |
14 |
10 |
3 |
||||
东非 |
6800年 |
12 |
31日 |
22 |
||||
安的列斯群岛东部 |
1920年 |
12 |
NA |
NA |
||||
中美洲 |
4630年 |
10 |
NA |
NA |
||||
密克罗尼西亚群岛 |
700年12 |
8 |
2 |
1 |
||||
北加勒比 |
9800年 |
5 |
4 |
3 |
||||
红海 |
17 640年 |
4 |
4 |
2 |
||||
西南太平洋岛屿 |
27日060年 |
3 |
10 |
8 |
||||
澳大利亚和巴布亚新几内亚 |
62 800 |
2 |
3 |
1 |
||||
波利尼西亚群岛 |
6733年 |
2 |
1 |
1 |
||||
夏威夷群岛 |
1180年 |
1 |
NA |
NA |
||||
总 |
284 803 |
20. |
16 |
6.4 |
||||
NA =不适用,因为没有损失记录是在1998年。 |
||||||||
1985 1990 1995 2000 2005
一年
图2最大年度热应力东加勒比海珊瑚礁作为加热程度月(月DHM°C)自1985年以来。DHM等于1个月ofSSTthatis1°C以上每月最大气候学的面积(改编自Ref。[19],版权(2007)国家科学院,美国)。
1985 1990 1995 2000 2005
一年
图2最大年度热应力东加勒比海珊瑚礁作为加热程度月(月DHM°C)自1985年以来。DHM等于1个月ofSSTthatis1°C以上每月最大气候学的面积(改编自Ref。[19],版权(2007)国家科学院,美国)。
3所示。相关的动物群的珊瑚礁
与珊瑚本身不同,相对较少的研究都集中在如何影响气候和全球变化巨大的生物多样性与珊瑚礁(大约四分之一的所有已知的海洋物种[13日30]),或许是由于物流建设长期数据集的困raybet雷竞技最新难和费用在这些生物。此外,研究鱼类种群变化及其直接猎物其他压力影响较大,尤其是钓鱼压力[13],使它更加难以确定气候变化的信号。raybet雷竞技最新然而,尽管有超过4000种鱼类与珊瑚礁[31]相关联,值得注意的是很少有研究解决气候变化对这一群体的影响。raybet雷竞技最新几乎没有电流的证据直接影响气温上升的珊瑚礁鱼,主要是因为缺乏热耐受[32]的数据,尽管一项研究在热带东太平洋[33]确定临界热Maxima 15鱼类的范围(34.7 - 40.8°C)高于记录温度(32°C)记录在1997年1998年厄尔尼诺事件。同样,缺乏珊瑚鱼类分布变化的证据,尽管被预测的气候影响评价[31]。raybet雷竞技最新这可能是由于缺乏除非珊瑚礁栖息地也扩展他们的范围[27]。
然而,有证据表明,珊瑚礁鱼可以影响更严重的间接影响大规模珊瑚白化(34 36)。在塞舌尔群岛珊瑚礁的研究在1997年1998年的厄尔尼诺现象[37],强烈的漂白导致减少鱼类丰度和组合内的转变从cor-allivores物种捕食无脊椎动物。鱼的大小结构也改变了与大型鱼类[38],可能时间上的滞后反应由于珊瑚结构复杂性影响鱼类的减少招聘,因此青少年的数量。然而,这个1997年1998事件的影响很小指出cryptobenthic多样性和丰富的鱼类在大堡礁[39],显示珊瑚礁鱼类可能会相对有弹性的短期扰动[39]只要礁结构持续[36]。
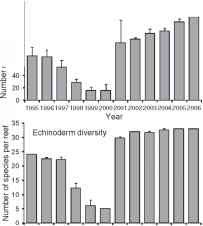
最广泛研究的影响气候事件对coral-associated无脊椎动物(> 500种)已经raybet雷竞技最新在巴伊亚,巴西、Kelmo和他的同事(40 42)。1997年1998年厄尔尼诺导致异常高温和减少通常的高浊度的区域允许更多的紫外线到达珊瑚礁。所有组研究(海绵除外)显示广泛的多样性减少多年之后厄尔尼诺现象(图3),包括当地的珊瑚灭绝一个物种(Porites astreoides);这是最有可能由于广泛的肿瘤肿瘤后的珊瑚紫外线的伤害[43]。大多数物种的密度也急剧减少(图3),只海胆Diadema antillarum显示一个投机取巧的应对不断变化的竞争条件和失踪类群[44]。这个数据集是引人注目的经济复苏组2001年之后,回到多样性,或超过水平之前,厄尔尼诺现象(图3)。礁组合显然已经从极端气候事件中恢复的能力,但前提是没有进一步的此类事件随后raybet雷竞技最新
4 O
16 7日14
12米
珊瑚密度
苔藓虫密度
1995199619971998199920002001 20022003200420052006
一年
棘皮动物密度
——1 |
莱托 |
J -。 |
I-IL。 |
||||||
杰。 |
我 |
||||||||
n ^猕 |
|||||||||
2006 2005 2002 2003 2004 20002001 19951996199719981999
一年
珊瑚的多样性
19951996199719981999 2000 19951996199719981999
一年
1995199619971998199920002001 20022003 2004 20022003
一年
苔藓虫多样性
1995199619971998199920002001 20022003200420052006
一年
棘皮动物密度
1995199619971998199920002001 20022003 2004 20022003
一年
苔藓虫多样性
19951996199719981999 2000 2001 19951996199719981999 2004 2005 2006
一年
2006 2005 2002 2003 2004 20002001 19951996199719981999
一年
19951996199719981999 2000 2001 19951996199719981999 2004 2005 2006
一年
图3三个主要群体的密度和多样性变化的无脊椎动物(珊瑚、苔藓虫、棘皮动物)在巴伊亚的补丁礁,巴西自1995年以来,表明1997年8厄尔尼诺的影响措施,加上标记康复三年后事件所有组除了珊瑚的多样性。水平的棘皮动物后复苏和苔藓虫多样性高于前厄尔尼诺期间。
S7.5
发生;在巴伊亚,没有主要的厄尔尼诺现象或漂白事件是明显的1998年之后。模型表明,ENSO事件在未来可能更频繁和严重的[8],这恢复珊瑚礁社区的能力可能受到气候变化。raybet雷竞技最新
4所示。结论
有清晰、明确的证据表明气候变化正在影响珊瑚礁系统[12],与特定担忧大规模珊瑚白化由raybet雷竞技最新于气温升高和随后的影响这对珊瑚的生存,因此相关的有机体。虽然珊瑚有一种天生的能力要适应这样的变化[45],正如过去[12],严重和常规的厄尔尼诺事件加上现代协同的影响其他人类活动,如钓鱼,污染(包括相对快速的海洋酸化[11])和旅游使珊瑚礁在世界上最脆弱的生态系统在当前场景未来的气候变化。raybet雷竞技最新
引用
1。c·帕尔玛,安。启生态。另一个星球。系统,37 (2006)637 669。
2。广义相对论沃尔特,大肠,p .传达门泽尔,c·帕尔玛T.J.C. Beebee, J.M. Fromentin, o·哥尔柏格赫伊,f . Bairlein自然416 (2002)389 395。
3所示。c·帕尔玛,水珠。改变生物13 (2007)1860 1872。
4所示。c·帕尔玛·g·尤伊,自然421 (2003)37 42。
5。A.L.佩里,P.J.低,jr埃利斯,J.D.雷诺兹,科学308 (2005)1912 1915。
6。中华民国霍尔布鲁克,R.J.施密特,j.s斯蒂芬斯,生态。:7 (1997)1299 1310。
7所示。C.D.G.哈雷,境休斯K.M. Hultgren,人矿工C.J.B.分拣器,C.S. Thornber, L.F. Rodriguez l . Tomanek S.L.威廉姆斯,生态。列托人。9 (2006)228 241。
8。a . Timmermann j . Oberhuber a·巴彻m .每m·拉蒂夫e . Roeckner自然398 (1999)694 697。
10。动向木匠,m . Abrar g . Aeby R.B. Aronson,美国银行,a·布鲁克纳奇,j .议会J.C. Delbeek, l . DeVantier G.J.埃德加,A.J.爱德华兹,d·芬纳古斯曼,
B.W. Hoeksema g·霍奇森,o .约翰W.Y. Licuanan,狭义相对论利文斯顿E.R. Lovell J.A.摩尔,交货单奥布拉,d . Ochavillo文学士学位Polidoro, W.F.普雷,m.c Quibilan (c . Reboton区域时间理查兹,公元罗杰斯,j . Sanciangco a·谢泼德c·谢泼德j·史密斯,斯图尔特,e . Turak J.E.N.贝隆,c·华莱士,大肠Weil, e .木材科学321 (2008)560 563。
11。o .哥尔柏格赫伊P.J. Mumby A.J. Hooten, R.S. Steneck p·格林菲尔德e·戈麦斯
C.D. Harvell,功率因数,A.J.爱德华兹,k .卡尔德拉n .伟达Eakin, r·伊格莱西亚斯普列托n . Muthiga r.h布拉德伯里,a . Dubi卷Hatziolos科学318 (2007)1737 1742。
12。t休斯A.H.贝尔德,湄Bellwood m·卡S.R. Connolly, c . Folke r . Grosberg o·哥尔柏格赫伊,J.B.C.杰克逊,j .形成J.M.湖,p .马歇尔m . Nystrom S.R.帕卢比,J.M. Pandolfi, b . Rosen, j .得科学301 (2003)929 933。
13。马丁Parry O.F. Canziani,摩根大通(J.P. Palutikof P.J.范德林登,石球汉森,十字章气候变化研究中raybet雷竞技最新,2007:影响,适应和脆弱性。第二工作组的贡献,政府间气候变化专门委员会第四次评估报告,剑桥大学出版社,剑桥,英国,2007年,页843 - 868。
14。J.B.C.杰克逊M.X.科比,w•h•伯杰·Bjorndal, l博茨,,B.J. Bourque r.h布拉德伯里,r·库克j . Erlandson J.A.埃斯蒂斯,t休斯,s . Kidwell C.B.兰格,H.S. Lenihan, J.M. Pandolfi, h彼得森,R.S. Steneck M.J. Tegner,水银血压计华纳,科学293 (2001)629 638。
15。摩根大通(J.P.威廉姆斯,贝聿铭象牙海岸,J.A.吉尔,j•萨瑟兰,境Watkinson、生态86 (2005)2055 2060。
16。公元巴顿,K.S.凯西,珊瑚礁24 (2005)536 554。
17所示。湄Bellwood, t休斯,c . Folke m . Nystrom自然429 (2004)827 833。
18岁。3月Freshw o·哥尔柏格赫伊。> 50 (1999)839 866。
19所示。唐纳,时任顶替克努森,m·奥本海默Proc。国家的。学会科学。美国104 (2007)5483 5488。
20.j•唐纳,时任Skirving,小,m·奥本海默哥尔柏格o·赫伊,水珠。改变生物11 (2005)2251 2265。
22。美国巴德梅尔,D.G. Fautin,生物科学43 (1993)320 326。
23。r . Berkelmans g . De 'ath s Kininmonth j•Skirving,珊瑚礁23 (2004)74 83。
24。c·威尔金森(主编),世界珊瑚礁的状态:2004年,1卷,目的,汤斯维尔,澳大利亚,2004年,p . 316。
25。R.B. Aronson, W.F.普雷西、硕士Toscano K.H. Koltes, 3月杂志。141 (2002)435 447。
26岁。k .罗伊·d·雅布隆斯基,J.W.情人节,生态。列托人。4 (2001)366 370。
27。W.F.普雷R.B. Aronson,前面。生态。环境。2 (2004)307 314。
28。助教加德纳,贝聿铭象牙海岸,J.A.吉尔,a·格兰特,境Watkinson、生态86 (2005)174 184。
29。n .伟达Proc。国家的。学会科学。美国98 (2001)5419 5425。
30.罗伯茨,C.J. McClean J.E.N.贝隆,摩根大通(J.P.霍金斯,广义相对论艾伦D.E.麦卡利斯特,C.G. Mittermeier, fw Schueler, m·斯伯丁f·威尔斯,c . Vynne结核病维尔纳,科学295 (2002)1280 1284。
31日。P.L.芒,他的琼斯,硕士,A.J.威廉姆斯,鱼鱼。9 (2008)261 285。
32。J.M. Roessig,伍德利,J.J.切赫,l·j·汉森,启鱼类生物。鱼。14 (2004)251 275。
33。c·莫拉自动跟踪Ospina, 3月杂志。139 (2001)765 769。
34。G.A. D.J.亭伯莱塔,3月生态。掠夺。Ser。245 (2002) 205 212。
35。K.C. Garpe S.A.S. Yahya,林达尔m.c欧曼(3月生态。掠夺。Ser。315 (2006) 237 247。
36。美国林达尔m.c欧曼(C.K. Schelten, 3月Pollut。公牛。42 (2001)127 131。
37岁。医学博士斯伯丁,ge贾维斯,3月Pollut。牛,44 (2002)309 321。
38。N.A.J. Graham,詹宁斯,威尔逊N.V.C. Polunin, j·罗宾逊,摩根大通(J.P.珠宝首饰,T.M.寒鸦,Conserv。杂志,21 (2007)1291 1300。
39岁。湄Bellwood, A.S.霍利,J.L. Ackerman m . Depczynski水珠。改变杂志。12 (2006)1587 1594。
40。f . Kelmo M.J. Attrill说M.B.琼斯,爬。26日》(2004)151 158。
41岁。f . Kelmo M.J. Attrill R.C.T.戈麦斯,M.B.琼斯、生物。Conserv。118 (2004) 609 617。
42。f . Kelmo M.J. Attrill说M.B.琼斯,珊瑚礁22 (2003)541 550。
43。f . Kelmo生态后果的1997 98厄尔尼诺南方涛动主要珊瑚礁社区从北部的巴伊亚,巴西、普利茅斯大学、普利茅斯,2002,第245页。
44岁。f . Kelmo Estuar M.J. Attrill说。海岸。架子上科学,73 (2007)243 248。
45岁。P.J. Edmunds, r•盖茨3月生态。掠夺。Ser。361 (2008) 307 310。
继续阅读:海洋生物多样性的变化作为气候变化的指标raybet雷竞技最新
这篇文章有用吗?
读者的问题
-
ALBERICA2个月前
- 回复
-
zewdi3个月前
- 回复
-
rahel1年前
- 回复