海洋生物远洋和浮游生态系统气候和全球变化的一个指标raybet雷竞技最新
马丁•爱德华兹
爵士Alister Hardy海洋科学基金会城堡山,锄头,普利茅斯PL1 2 pb,联合王国
普利茅斯大学海洋研究所,德雷克马戏团,普利茅斯PL4 8 aa,联合王国
1。远洋和浮游
生态系统
1.1。远洋的敏感性和浮游生态系统对气候和全球变化raybet雷竞技最新
1.2。海洋和陆地生物对气候和全球变化的反应raybet雷竞技最新
1.3。海洋酸化和其他人为对远洋和浮游生态系统的影响
2。观察对远洋和浮游生态系统的影响
2.1。生物地理学的变化和向北转移
2.2。生命周期事件和远洋生物气候学
2.3。浮游生物丰度和远洋的生产力
2.4。深海生物多样性和物种入侵
3所示。结论和摘要的关键指标参考
1。远洋和浮游生态系统
海洋深海区是地球上最大的生态系统占据71%的行星表面和整体地球生物圈的重要组成部分。因此,深海生态系统发挥着基础性的作用在调节全球环境通过其监管对地球的影响
raybet雷竞技最新:观察到的气候变化对地球的影响
版权©2009爱思唯尔帐面价值保留所有权利的任何形式的复制。
raybet雷竞技最新气候及其在生物地球化学循环中的作用。变化引起的气候变暖对海洋远洋社区可能会增加生态结构和功能有重要的影响从而导致地球的气候系统上重要的反馈。raybet雷竞技最新
本章将主要集中在上层的区域生物生产、生物地球化学循环和海洋食物网是由浮游生物居住。除了讨论高等营养生物气候的影响,尤其是远洋鱼,整个这一章的重点集中在浮游社raybet雷竞技最新区。更具体地说,本章将重点观测证据从当代浮游生物指标在过去的数十年时间,而不是古浮游指标。海洋的自由浮动的光合作用的生活(藻类浮游植物、细菌和其他光合作用原生生物),底部的海洋食物网,为动物提供食物浮游生物(浮游动物),反过来,为许多其他海洋生物提供食物从微观到鲸鱼。深海生态系统的承载能力的大小的鱼资源,招聘个股以及丰富的海洋野生动物(例如,海鸟和海洋哺乳动物)是高度依赖变化丰富,季节性的时机和浮游生物的成分。
浮游植物也约占全球初级生产总数的一半,通过生物在气候变化中扮演着重要的角色raybet雷竞技最新地球化学循环和出口的温室气体的深海碳封存在所谓的“生物泵”。Phyto-plankton已经因此有助于减轻一些观察到的高二氧化碳对气候造成的影响在过去的200年海洋占用^ 40%的人为二氧raybet雷竞技最新化碳[1]。对地球气候反馈机制,推测这些生物泵将在变暖的世界由于低效率的浮游植物组成的变化有利于小raybet雷竞技最新鞭毛虫[2]和更少的总体营养混合由于分层的增加(见1.1节)。它也预测,气温将世界上生产和呼吸代谢平衡对增加呼吸的海洋从而减少海洋捕获二氧化碳的能力[3]。除了发挥根本性作用在地球气候系统和海洋食物网,浮游生物也高度敏感的现代和古环境变化的指标,并提供快速的信息raybet雷竞技最新在我们的海洋的生态健康。有一些证据表明,浮游生物指标更敏感比环境变量本身,可以放大微弱的环境信号由于其非线性反应[4]。浮游生物物种,由其非生物信封,在同时有能力代表了一个完整的生态影响,化学和物理变量。
1.1。远洋的敏感性和浮游生态系统对气候和全球变化raybet雷竞技最新
温度是一个海洋生态系统的关键驱动因素,特别是,它对远洋影响人口迅速体现7 [5]。这是当超过99%的远洋,不足为奇浮游生物ectothermal使它们的波动高度敏感温度[8]。浮游的速度反应主要是由于他们的短生命周期及其被动应对平流变化。例如,phyto-plankton修复尽可能多的二氧化碳每年所有陆生植物但由于单细胞他们代表在任何时候只有1%的地球生物。这意味着在世界上的海洋是巨大的周转率和平均全球浮游植物的人口消耗在天周[9]。这一切使浮游生物紧密耦合波动的海洋环境和营养等环境变化的高度敏感指标可用性,洋流的变化和气候变raybet雷竞技最新化。
在海洋环境的影响短期气候变化、年际变化更高营养级如海鸟和鲸的数量在一定程度上可以有所缓冲由于其较长的生raybet雷竞技最新命周期。从长远来看,他们的接受能力大地理迁移也可能帮助他们减轻一些全球变化的影响;然而,这个假设没有被调查。在这两种情况下,这并不适用于浮游生物。生物学上来说,温度的变化对许多生理过程有直接影响(如氧代谢,成人死亡率、复制、呼吸、生殖发展,等等)和控制几乎所有生命过程从分子到细胞整个区域生态系统水平和生物地理学省份。生态而言,温度也调节直接或间接地物种相互作用(如竞争、捕食者猎物的相互作用和食物链结构),最终,温度的变化会导致对生物多样性的影响,大小整个深海生态系统的结构和功能(10、11)。
,温度直接影响在许多生物和生态特征,它还修改了海洋环境通过影响海洋环流和提高潜的稳定性,因此营养的可用性。营养素的数量可以在地表水直接决定浮游植物生长,浮游生物的大小的关键因素,社区和食物网结构。营养方面的可用性,表面变暖层增加水体的稳定性,提高分层和需要更多的能量混合深,营养丰富的水域在表面层次。特别是温暖的冬天也会限制的程度深对流混合,从而限制以下的营养补充必要的春天浮游植物爆发。总之,气候变暖的表层水会增加表层和底层之间的密度差营养丰富的水域。营养原则之一的可用性(硝酸盐),因此限制了浮游植物生长被发现对全球温度负相关(12、13)。同样,全球卫星提取叶绿素数据分析显示了强烈的逆关系海表温度(SST)和叶绿素浓度[9]。此外,其他非生物变量如氧浓度(重要器官大小和代谢[14]),硝酸代谢[15]和粘度的海水(浮游生物重要的维护浮力)也直接与温度有关。与陆地环境中,降水中发挥着关键作用,化学和海洋上层温度制度在开放的海洋和它的顺向生物成分是不可避免地纠缠在一起。
1.2。海洋和陆地生物对气候和全球变化的反应raybet雷竞技最新
许多浮游生物生活在狭窄的温度范围(stenothermal)和经常进行更迅速观察到由于温度改变,生物地理学和物候(10、11),相比陆地同行[16]。除了这和浮游生物有更短的生命周期,这一事实已经上面提到的,有许多不同的原因的速度应对气候和全球变化的深海生物不同于陆地的生物。raybet雷竞技最新的主要原因是,第一,由于水在海洋开放系统的高比热许多浮游生物主要是缓冲极端每日和季节性的温度波动。每日和季节性温度变化因此少变量相比陆地域允许海洋物种成为他们最佳热牢牢地嵌在信封。其次,与陆地环境中,许多浮游生物可以快速跟踪进化生物气候学的信封被很大程度上免费的地理障碍阻碍他们的传播范围,不需要大量的能量消耗,主要是被动地流水。洋流,因此,提供了一个理想的远距离传播机制,这似乎就是为什么一个巨大的海洋生物已经进化至少部分浮游实体的生命周期。第三,许多陆地生物地理和生态受栖息地类型主要是由营养成分。在陆地系统中,这些营养类型的发展尤为缓慢移动(例如,森林生态系统),因此生物栖息地将依赖于此限制的地理分布。情况不是这样的浮游植物极短的生命周期相比,允许快速时空传播浮游食草动物和相关社区。此外,独特地陆地的存在人为压力如栖息地的分裂和栖息地的丧失,这显然限制了生物的地理分布在陆地环境中,似乎缺席大洋系统[17]。
1.3。海洋酸化和其他人为对远洋和浮游生态系统的影响
在温度、光和营养可能是最重要的物理变量构建海洋生态系统,深海领域也将不得不面对,除了全球气候变化,人为二氧化碳的影响直接影响海洋pH值[18]。raybet雷竞技最新证据收集和模仿迄今为止表明上升的二氧化碳导致了化学海洋的变化导致海洋酸化。海洋酸化有潜力影响钙化的过程,因此某些浮游生物(如颗石藻,foraminif-era,远洋软体动物)可能特别容易受到未来的二氧化碳排放。除了气候变暖,海洋潜在raybet雷竞技最新的化学变化及其对海洋生物的影响可能会进一步减少海洋从大气中吸收更多的二氧化碳的能力,进而可能影响全球变暖的速度和规模(见21章)。其他人为驱动力变化的手术在深海生态系统主要是对修改海洋过度捕捞和其影响远洋食物网[19],在沿海地区(见第14章)和养分输入从陆地来源导致在某些情况下提高生物产量和有害藻华(赤潮)和其他通用化学和无机污染物。大气中人为的影响氮在开放海域只有最近调查,但也可能扮演了一个重要的角色在年度新的海洋生物生产[20]。
2。观察对远洋和浮游生态系统的影响
有大量的观测证据表明,许多深海生态系统,物理和生物都是应对区域气候的变化主要是由于气候变暖造成的空气和SST和在较小程度上的降水机制和修改raybet雷竞技最新风的模式。上升的生物表现SST有不同的生物地理学的形式,物候、生物多样性、生理、物种丰度变化和整个生态体系转变。任何观察海洋环境的变化与气候变化有关,然而,应该考虑自然变化的背景下,在不同的空间和时间尺度。raybet雷竞技最新最近,长期十年观察性研究都集中在已知的自然气候振荡模式类似的时间尺度,如厄尔尼诺-南方涛动(ENSO)
太平洋和北大西洋涛动(NAO)在北大西洋深海生态系统变化的关系(见评论(21、22))。许多生物反应的观察与气温升高有关。然而,近似的影响气候变化嵌入在自然模式的可变性,尤其是数十年振荡像大西洋数十raybet雷竞技最新年震荡(AMO)[23],是极其困难的,因此观察到的证据浮游直接归因于人为气候变化与全球变化必须接受一定程度的科学谨慎。
证据观察远洋变化也偏向地区,特别是欧洲和北美周边海域,有某种形式的生物监测在持续长时间。除了一些重要的长期沿海研究站抽样浮游生物(例如,Helgoland道路时序北海南部[24])只有少数长期开阔的海洋生物调查样本。因此一些观察到的变化的最有力的证据发现开放的海洋生态系统来自北大西洋的一个广泛的空间和长期生物调查的形式存在的连续浮游生物记录器(CPR)调查。CPR的调查已经在北海和北大西洋自1931年以来,系统抽样500浮游类群从北大西洋的主要地区每月决议[25]。重要数十年来自太平洋的证据主要来自加州的合作海洋渔业调查(CalCOFI)调查操作自1949年以来,加州海岸。
2.1。生物地理学的变化和向北转移
一些大型生物地理学的最有力的证据的变化在我们的海洋来自CPR的调查。在地理上的一项研究包括在50年的时间里,整个东北大西洋Beaugrand et al。[10]显示快速向北运动的生物多样性的一个关键的浮游动物组(calanoid桡足类)。在过去的50年里一直向北运动的温暖海水浮游生物10°纬度在东北大西洋和类似的撤退的北部寒冷的水中浮游生物(意味着向极运动之间的200和250公里每十年)(图1)。这种地理运动更明显比任何记录地面研究中,主要是由于平流过程,特别是北沿着崖边当前运行的欧洲北部大陆架。浮游生物的快速运动沿着大陆架向北只是见过,在更深的水变暖更快。沿着书架,浮游生物是upwelled从这个更深的水出现在水面浮游生物群落。因此浮游生物已经向北10°纬度主要通过深水平流过程未见的表面等温线的运动。在其他领域在北方
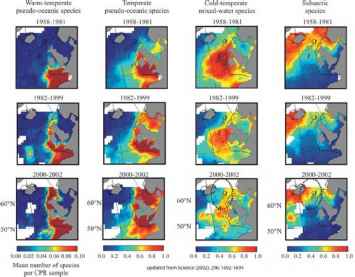
图1中四种不同的浮游生物的地理分布的变化组合在数十年的时间里。有温暖的温带物种的快速向北运动,随后下降子北极物种超过40年。尤其是快速运动是沿着欧洲大陆架观察。来自连续浮游生物记录器的数据调查。更新来自Ref。[76]。
图1中四种不同的浮游生物的地理分布的变化组合在数十年的时间里。有温暖的温带物种的快速向北运动,随后下降子北极物种超过40年。尤其是快速运动是沿着欧洲大陆架观察。来自连续浮游生物记录器的数据调查。更新来自Ref。[76]。
东大西洋之间的浮游生物变化更温和的和多样的90和200公里每十年,仍然比任何其他记录地面研究整合的平均每十年6公里[16]。类似于北大西洋,在东北太平洋有一般的频率增加物种南部[26]向北移动。有趣的是,在西北大西洋深海生物一直向南移动[27]。这最初似乎矛盾的一般思维同质全球气候变暖的世界各地的海洋中。raybet雷竞技最新然而,这场运动的加强与拉布拉多寒流向南传播冷的水在过去的十年里携带着远洋生物冷水亲和力南至乔治银行。
这些大规模的生物地理学中观察到的浮游生物变化也看到平行的鱼类分布的纬向运动30 [28]。北方的地理范围扩展或鱼类种群的地理分布的变化最近欧洲大陆架海域和记录欧洲大陆架边缘33 [31]。类似于浮游生物,最大的鱼类向北运动沿着欧洲大陆架也被观察到。这些地理运动与区域气候变暖,主要是与鱼类有南方的北方地理运动生物地理学紧密联系。raybet雷竞技最新这些包括远洋鱼类的运动如沙丁鱼和凤尾鱼向北在北海和红鲻鱼和低音扩展他们的范围向北挪威西部(31、32)。新记录也被观察到在过去十年的地中海和非洲西北部南海岸的物种葡萄牙[32]。淡化的冷却和西北大西洋在过去的十年里已经有一个相反的效果类似于浮游生物模式,和一些南部的底栖鱼物种移动地理分布[34]。北端的范围扩展的远洋鱼类也有北方白令海地区与地区气候变暖[35]。raybet雷竞技最新raybet雷竞技最新气候变化和区域气候变暖也与海洋的地理范围的变化相关疾病[36]。新的疾病通常出现在主机或已知病原体的变化范围。 For example, over the past few decades pathogens detrimental to oysters have spread from the mid-Atlantic states into New England [36]. In comparison to terrestrial systems, epidemics of marine pathogens can spread at extremely rapid rates [37].
同样值得注意的是鱼与分布在北海边界北部向北移的速度比陆地物种快3倍[16,30]。有史以来最大的生物地理学的变化观察鱼类的急剧增加和随后的地理分布向北的蛇海龙(Entelurus aequreus)。一旦主要局限于南部和西部的不列颠群岛在2003年之前,它现在可以发现巴伦支海和北Spitzbergen[38]。虽然现在讨论描述了表面上层的生物地理变化值得记住的深海环境的三维特性。最近的研究观察到的不仅仅是鱼类生物地理学的变化也改变鱼类深度(向更深的水域)在应对气候变暖[39]。raybet雷竞技最新这种变化可以被视为与陆地生物的上升高度的运动类似的高山环境。所有这些研究强调的一致性远洋生物发生大规模分布变化以应对hydro-climatic可变性。
2.2。生命周期事件和远洋生物气候学
物候学或重复季节性生命周期事件,如年度迁移或产卵,气候变暖的一个敏感指标。raybet雷竞技最新这是因为许多陆地和海洋生物,除了光周期,是依赖于温度触发季节性行为。在陆地领域,生物气候学等事件鸟迁徙,卵生,蝴蝶出现和开花的植物都得到反应温和的春天早些时候天气[16]。在远洋物候应对气候变暖,还发现了许多浮游生物类群在他们前进raybet雷竞技最新季节性周期[11]。在某些情况下,转变季节性周期超过6周检测,又一个更大的转变比观察基于地面观测。总结地面物候学的研究使用超过172种植物、鸟类、昆虫和两栖动物、帕玛森&约克[16]计算平均2.3 d的物候变化。人们认为温带深海环境中特别容易受到气候变暖导致的物候变化,因为招聘的成功率较高的营养级高度依赖于同步脉冲浮游生产[11]。此外在海洋环境中,同样重要的是,是区域气候变暖的响应之间的各种不同的官能团和营养水平,导致营养级之间的时间不匹配(图2)。例如,在春天开花季节性时间在五几十年一直保持相对稳定(主要是raybet雷竞技最新由于光线的限制,光周期而不是温度指示季节性[11,40])许多浮游动物有机体以及小鱼已经迅速向前的季节性周期。
这些变化,在北海,有潜力成为损害商业鱼类通过营养不匹配。例如,在北海地区气候变暖已经影响到鳕鱼招聘通raybet雷竞技最新过改变底部的食物链[41]。鳕鱼,和其他鱼类一样,高度依赖于食品在深海浮游幼虫阶段的可用性。关键浮游组合的变化和物候学,显著相关的变暖北海在过去的几十年里,已经导致了一个可怜的鳕鱼幼虫食物环境,因此最终下降总体招聘成功。浮游生物群落的快速变化观察到在过去的几十年里在北大西洋和欧洲地区海域,与地区气候变化,对其他营养水平和生物地球化学过程有巨大的影响。raybet雷竞技最新同样,其他远洋生物气候学的变化被观察到在北海[24],地中海[42]和太平洋(43、44)。
2.3。浮游生物丰度和远洋的生产力
当代的观测卫星原地混合海洋叶绿素记录表明,全球海洋净初级生产量下降在过去的十年里[9]。虽然这个时间序列长度只有10年它表现出强烈的负面关系主要生产和SST和证据之间的紧密耦合关系海洋生产力和在全球范围内气候变化。raybet雷竞技最新在北大西洋和经过数十年时间,浮游植物和浮游动物物种的变化和社区与北半球温度相关(3)趋势和变化NAO指数。这些包括物种分布的变化和丰富,亚热带物种的出现
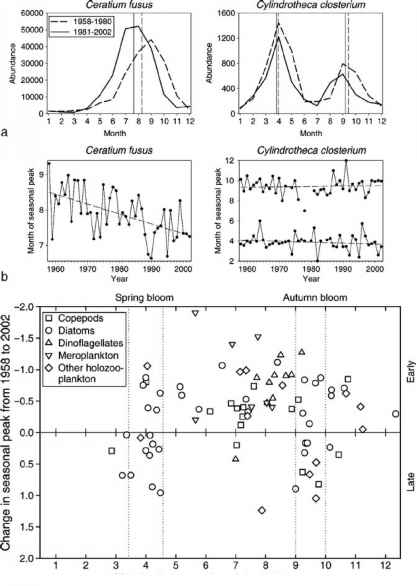
图2 (a)季节性周期两个浮游植物时间1958年2002年1980年和1981年:腰鞭毛虫的角藻fusus和硅藻Cylindrotheca梭形藻属。(b)国际年度季节性高峰的杂文集性上面的两个物种从1958年到2002年。(c)的变化季节性高峰的时间(几个月)66类群45(年)时期从1958年到2002年策划针对季节性的峰值的时间在1958年。对于每一个分类单元,(b)的线性回归是用来估计季节性高峰在1958年和2002年之间的差异。- 1958年和2002年之间的差异表明季节性周期越来越早。标准线性回归被认为是适当的,因为是最小的自相关(由德宾沃森统计)物候变化时间序列。Ref。[11]。
3 4 5 6 7 8 9 c 1958年月的季节性高峰
图2 (a)季节性周期两个浮游植物时间1958年2002年1980年和1981年:腰鞭毛虫的角藻fusus和硅藻Cylindrotheca梭形藻属。(b)国际年度季节性高峰的杂文集性上面的两个物种从1958年到2002年。(c)的变化季节性高峰的时间(几个月)66类群45(年)时期从1958年到2002年策划针对季节性的峰值的时间在1958年。对于每一个分类单元,(b)的线性回归是用来估计季节性高峰在1958年和2002年之间的差异。- 1958年和2002年之间的差异表明季节性周期越来越早。标准线性回归被认为是适当的,因为是最小的自相关(由德宾沃森统计)物候变化时间序列。Ref。[11]。
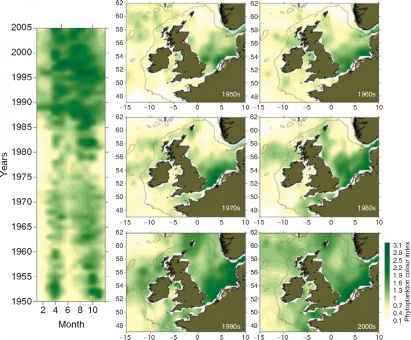
图3的空间时间地图的变化丰富的浮游植物色彩(浮游植物总生物量指数)东北大西洋平均每十年从1950年代到现在。等高线图显示了从1950年到2005年月度平均值平均浮游植物色彩的北海。大量增加浮游植物的颜色的观察到1980年代末,一直以来。颜色的增加与政权有关北海的转变。更新来自Ref。[47]。
图3的空间时间地图的变化丰富的浮游植物色彩(浮游植物总生物量指数)东北大西洋平均每十年从1950年代到现在。等高线图显示了从1950年到2005年月度平均值平均浮游植物色彩的北海。大量增加浮游植物的颜色的观察到1980年代末,一直以来。颜色的增加与政权有关北海的转变。更新来自Ref。[47]。
在温带水域,浮游植物总生物量的变化和季节性长度(图3),生态系统功能和生产力的变化北大西洋(10 11 45 52)。在北海浮游植物总生物量的增加与增加有关小鞭毛虫支持更多的温暖和分层条件[46、47]。在整个东北大西洋有浮游植物生物量随着温度的增加在寒冷地区但浮游植物生物量下降[53]在温暖的地区。据推测,这是一个之间的权衡增加浮游植物代谢率引起的温度凉爽地区但减少养分供应在温暖的地区。在北海地区气候变暖raybet雷竞技最新也伴随着某些赤潮的增加在某些领域的北海[54]。最近链接之间建立了浮游生物的变化在北海sandeels和最终的海鸟繁殖成功(包括四个营养级)[55]。在北方
海,以前占主导地位的人口和生态重要的浮游动物物种(冷水物种Calanus finmarchicus)自1960年代以来在生物量下降70% [56]。物种温暖海水亲和力正在向北来取代这个物种丰富但这些物种不是数值或营养有利于营养水平较高。这不可避免的重要影响北海的整体承载能力的生态系统。
生态变化发生在北海自1980年代末(主要由温度改变政权,更温暖的冬天)也被记录为波罗的海浮游动物和鱼类资源[57,58]。相关的变化,在这些北欧海域发生了足够的和持续的被称为“政权转移”[59]。同样在地中海,浮游动物群落也与地区变暖,NAO指数[60]。所有这些观察到的变化似乎是密切相关的气候驱动海洋温度波动的影响。raybet雷竞技最新间接的逐步淡化拉布拉多海地区,由于气候变暖海洋和淡水输入的增加从融化的冰,导致越来越丰富,花朵和季节性的变化周期的鞭毛藻类由于增加稳定的潜[61]。raybet雷竞技最新同样,增加coccolithophore花朵在巴伦支海和赤潮在北海与消极盐度异常和气温升高导致增加分层[62]。
在本格拉上升流系统在南大西洋,长期趋势丰富和沿海浮游动物的群落结构与大规模的气候影响[63]。同样,mesozoo-plankton丰度的变化也被大规模的加州的气候影响上升流有关系统[64]。raybet雷竞技最新逐步变暖在南大洋与磷虾下降有关[65]和一个相关的人口规模的下降许多海鸟和海豹监控几个繁殖地点(66、67)。在南大洋长期下降的磷虾股票与变化在冬季冰程度反过来一直与气候变暖有关[65]。大量的磷虾的变化对南大洋食物链产生深远的影响。南大洋的进步变暖也与人口规模的下降许多海鸟和海豹监控几个繁殖地[67]。最近调查的浮游有孔虫从沉积物包括过去1400显示异常群落结构的变化在过去的几十年里。研究表明,海洋变暖已经超过了自然变化的范围[68]。最近的重大生态系统转变在北部白令海归因于区域气候变暖,北极涛动的趋势[35]。raybet雷竞技最新年代际变化与气候变化相关的浮游动物在西方潜艇[69]中也发现了北极北太平洋和日本/东海[70]。
许多丰富的海洋经济鱼类资源变化被观察到在过去的几十年里在大西洋和太平洋,但很难分开,在人口密度的变化和招聘方面,区域气候效应从直接人为影响喜欢钓鱼。raybet雷竞技最新地理范围扩展前面提到的或鱼类种群的地理分布的变化,然而,可以更自信地与hydro-climatic变异和区域气候变暖。raybet雷竞技最新类似于观察到的变化在海洋浮游系统许多远洋鱼类种群的长期变化与ENSO等自然气候振荡模式和太平洋年代际振荡(PDO)在太平洋和北大西洋NAO(见评论:[5,21日,22日,26])。例如,海温由NAO波动的变化与波动鳕鱼招聘了拉布拉多和纽芬兰和巴伦支海[71]。人口的鲱鱼、沙丁鱼、鲑鱼和金枪鱼也被有关NAO指数波动(5,45岁)。温暖的事件与厄尔尼诺事件和气候引起的生态政权转移在太平洋中断相关的许多商业渔业[72]2raybet雷竞技最新1日26日。这些变化突出鱼类对环境变化的敏感性。直接证据的生物气候变化是人类活动的影响,然而,很难辨别由于自然变异的背景在各种空间和时间尺度上,特别是气候自然raybet雷竞技最新振荡。最近的一项研究基于50幼虫鱼CalCOFI时间序列显示,利用鱼类更容易受到气候变化的影响比non-exploited物种。raybet雷竞技最新作者建议,增强利用物种对环境变化的反应是由于减少空间异质性fishery-induced年龄引起的截断和地理分布的限制,同时捕捞压力[73]。
2.4。深海生物多样性和物种入侵
在海洋盆地规模的深海生物多样性研究浮游动物桡足类相关温度和气候变暖的增加在过去的几十年里一直是紧随其后的是多样性的增加(74 76)。特别是,增加多样性被认为当一个以前低多样性系统像北极和cold-boreal省份进行长期变暖事件。远洋的总体多样性模式生物,峰值之间20°和30°北或南,跟随世界上温度梯度的海洋[77]。同样,浮游植物显示温度之间的关系和浮游植物多样性与社区有一个更高的多样性,但整体规模较小的颗粒组和一个更复杂的食物网结构(即。,microbial-based与温暖更多的分层环境中基于硅藻的生产)。raybet雷竞技最新气候变暖将因此增加浮游生物多样性在寒冷地区的海洋温度等温线向极的转变。然而,温度和表层鱼类多样性之间的关系要复杂的多由于过度捕捞等人为压力显然中扮演重要角色的多样性模式[19](见第14章)。
raybet雷竞技最新气候变暖将开辟新的热定义栖息地此前曾否认本土物种(例如,亚热带物种在北海)和入侵物种允许他们建立可行的人口的地区曾经是环境不合适。除了这些热边界限制逐步向极和在某些情况下扩大移动,快速气候变化观测北极可能有更大的影响建立外来物种入侵和生物多样性的北大西洋。raybet雷竞技最新夏天的厚度和面积覆盖在北极冰已经融化以越来越快的速度在过去二十年中;2007年9月达到有记录以来的最低程度。在春天异常巨大冰后空闲时间1998年大量的太平洋硅藻Neodenticula seminae CPR调查的样本中发现了北大西洋的拉布拉多海。n seminae是一个丰富的浮游植物的近极的北太平洋,有一个定义良好的古历史根据深海内核。根据古证据和现代表面采样在北大西洋的自1948年以来,这是第一个记录这个物种在北大西洋的再现至少800 000。n . seminae在北大西洋,及其随后向南和向东蔓延到其他地区在北大西洋,经过这么长时间的差距,可能是一个指标的规模和速度的变化发生在北极和北大西洋海洋气候变暖的结果[78]。raybet雷竞技最新硅藻物种可能本身可能是第一个的证据trans-Arctic迁移在现代,预示着一个新的生物进入北大西洋的潜在泛滥。这样一个改变功能的后果,和生物多样性的北极气候反馈系统是目前未知。
3所示。关键指标的结论和总结
指一些强调集体综述表明,有大量的观测证据表明许多深海生态系统,物理和生物都是应对区域气候变化引起的气候变暖主要海温、洋流变化和修改在较小程度上的降水机制和风模式。raybet雷竞技最新气候变化的生物表现迅速采取生物地理学的形式,物候、生物多样性、生理、物种丰度的变化,社会结构变化和整个生态体系转变。一些最令人信服的证据的生物反应的区域气候变化来自底部的海洋食物网特别是远洋浮游植物和浮游动物的社区。raybet雷竞技最新与气候变暖有关的其他反应更高营养级也间接与浮游生物的变化和暗示自下而上的海raybet雷竞技最新洋深海环境的控制。因此假定种群应对气候的方式之一是在一定程度上取决于食物网结构的变化,人口是嵌入式,捕食者和猎物之间的同步(匹配不匹配)起着重要的作用。raybet雷竞技最新
在物种水平,第一批气候变暖的后果和全球变化通常出现在一个物种物候学(即。raybet雷竞技最新、时间年生命周期事件发生)和物种的地理分布响应。这主要是因为温度不断影响物种的生命周期和自然的人口将会反应随着时间的推移,提供它不是生物的抑制或空间限制,其最佳位置在其生物气候学的信封。这是否是在颞利基在季节性演替(观察物候反应)或在其整体biogeograph-ical分布(观察人口地理运动)。这些生理变化以及这些变化观察到生物多样性和浮游丰度和生产率可能是关键指标标志着大规模的变化发生在我们的全球海洋气候和全球变化的结果。raybet雷竞技最新
总结观察到的个案研究中得到,特别是突出综述远洋和浮游响应的速度,无论是biogeogra-phically或物候对气候变暖,全球变化与陆地同行相比。raybet雷竞技最新例如,浮游生物变化的200公里每十年[10]曾被观察到在大西洋东北部与整合相比陆地平均每十年6公里[16]。同样,改变生物气候学的6周已观察到深海生态系统[11]相比,平均物候的变化2.3集体观察172种植物、鸟类、昆虫和两栖动物[16]。无数的陆地和海洋领域之间的差异(见1.2节浮游的速度反应主要是由于他们的生命周期短,主要是被动应对平流变化。这些变化突出综述未来将继续遵循目前气候变暖的预测。raybet雷竞技最新因此认为目前观察到的和未来气候变暖很可能会继续改变中小学浮游的地理分布生产[53],影响海洋生态系统服务,如氧气生产、固碳和生物地球化学循环和放置额外的压力已经枯竭的鱼类和哺乳动物种群。
对地球气候反馈机制而言,人们认为这些生物泵将低效率在一个变暖的世界由于浮游植物组成的变raybet雷竞技最新化(植物区系变化)和更少的总体营养混合(减少批量属性)由于增加世界上的海洋的分层。特别是,这将影响大面积的热带海洋永久分层[9]。也有强烈的负面关系海洋生产力和SST(通过养分有效性有关)在全球范围内(9、12、13)。尽管气候变化及其空raybet雷竞技最新间异构影响表面风,风力强度、上升流和深-水混合让许多区域预测与不确定性困扰。同样值得注意的是,北半球的深海生物的潜在栖息地扩张由于北极冰层的融化将严格限制光口述季节性限制浮游植物生产。然而,许多这些场景仍在初期阶段,而这是相对简单的预测改变海洋物理气候强迫下,理解生物反应由于经常底层生物群落的复杂性及其非线性响应环境变化使得预测植物区系变化充满了不确定性。raybet雷竞技最新调查的重要性生物固氮作用和甲基(DMS)的生产某些浮游植物目前需要了解生物增加了氮循环的分层的后果(79、80)和生物反馈(81、82)。
生态演讲和在全球范围,浮游生物和深海生态系统作为一个隐喻集体实体,可能是一些最敏感的生物对环境变化和最重要的一个地球上生物群落。他们负责绝大多数的海洋生物燃料生产海洋食物网和养分循环以及导致世界上大约一半的氧气生产和碳封存。实际上,所有的生物观察强调综述源于财政脆弱的数十年监测项目。未来这些生态系统的生物监测,通过一个集成的和持续的观察方法,将在理解基本的持续影响气候和全球变化在我们的行星系统。raybet雷竞技最新这反过来可能允许我们通过国际合作来减轻和自适应管理他们的一些更多的有害影响。
引用
1。卡尔德拉,右眼齐伯,J.C. Zachos共同断定k, t . Tyrrell科学321 (2008)51 52。
2。l . Bopp o . Aumont p . Cadule s Alvain m . Gehlen地球物理学。卷,32(2005)货号。L19606。
3所示。a·洛佩兹乌鲁蒂亚e·圣·马丁r。哈里斯,x Irigoien, Proc。国家的。学会科学。美国103 (2006)8739 8744。
4所示。艾伦A.H.泰勒系统,公共广播克拉克,自然416 (2002)629 632。
5。K.F.时候,a . Belgrano (a .哈C.A.M.爱德华兹,h格林,g . Ottersen A.J.潘兴,h·沃克,地球物理学。专著134 (2003)211 243。
6。g . Beaugrand f . Ibanez说j·林德利3月生态。掠夺。Ser。219 (2001) 205 219。
7所示。g . Beaugrand pc里德,水珠。改变杂志。9 (2003)801 817。
8。. sib d·阿特金森,趋势生态。另一个星球,12 (1997)235 239。
9。R.T. M.J. Behrenfeld说奥马利,D.A. Siegel C.R.麦克莱恩,J.L. Sarmiento G.C.费尔德曼A.J. Milligan P.G.法尔.勒特里尔,静电的老板,自然444 (2006)752 755。
10。g . Beaugrand pc里德和f . Ibanez说j·林德利m·爱德华兹,科学296 (2002)1692 1694。
11。m·爱德华兹A.J.理查森,自然430 (2004)881 884。
12。d . Kamykowski中华民国Zentara、深海研究》(我Oceanogr。> Pap) 33 (1986) 89 105。
13。d . Kamykowski中华民国Zentara、深海研究》(我Oceanogr。> Pap) 52 (2005) 1719 1744。
14。H.O. Portner, r . Knust科学315 (2007)95 97。
15。J.A. berg, D.E. Varela P.J.哈里森,3月生态。掠夺。Ser。225 (2002) 139 146。
16。c·帕尔玛·g·尤伊,自然421 (2003)37 42。
17所示。l .汉娜式样洛夫乔伊,萨达姆政权施耐德:式样洛夫乔伊,l·汉纳(Eds),气候变化和生物多样性,耶鲁大学出版社,纽黑文,CT, 2005年,页3 14。raybet雷竞技最新
18岁。C.L. Sabine, R.A.费利,k . Lee, w . Berelson j .形成V.J. Fabry, F.J. Millero,科学305 (2004)362 366。
19所示。蠕虫,E.B.巴比尔:博蒙特,J.E.达菲,c . Folke B.S. Halpern J.B.C.杰克逊,香港Lotze, f . Micheli S.R.帕卢比,e·萨拉·阿兹J.J. Stachowicz, r·沃森科学314 (2006)787 790。
20.R.A.首领,j . LaRoche k·阿尔提耶里,投资者Arrigo,由拉贝克D.G.卡彭,美国康奈尔大学,f . Dentener j . Galloway R.S. Ganeshram R.J. Geider, t . Jickells M.M. kuyper, r . Langlois注:丽丝,克里,J.J.船帆,摩尔,s . Nickovic a . Oschlies彼得森t . j .普洛斯彼罗r . schlitz s Seitzinger L.L.索伦森,m . Uematsu o . Ulloa m·沃斯b . Ward, l .萨莫拉科学320 (2008)893 897。
21。j .陆路s Rodionov s Minobe n键,掠夺。Oceanogr。77 (2008) 92 102。
22。北卡罗来纳州Stenseth, g . Ottersen J.W. Hurrell, a . Mysterud m·利马K.S. Chan净收益Yoccoz, b . Aadlandsvik Proc。r . Soc。Lond。爵士。B:生物。科学,270 (2003)2087 2096。
23。R.T. Sutton D.L.R. Hodson,科学309 (2005)115 118。
24。w·Greve s Prinage h . Zidowitz j . Nast f . reiner冰j . 3月Sci 62 (2005) 1216 1223。
25。pc里德,J.M.科尔布鲁克J.B.L.马修斯j·艾肯,掠夺。Oceanogr 58 (2003) 117 173。
26岁。正当麦高文,湄Cayan L.M.多尔曼,科学281 (1998)210 217。
27。D.G.约翰·m·爱德华兹,板条,时任即可。j .鱼。Aquat。Sci, 58 (2001) 2121 2124。
28。j . c . Quero m·h·杜在j . j . Vayne Oceanol。Acta 21 (1998) 345 351。
29。k .打烙印,布罗姆·g·博尔赫斯第一,k . Erzini g·亨德森,开国元勋之一B.R.安贝德卡对MacKensie, h·门德斯,j·里贝罗A.M.P.桑托斯r . Toresen冰3月科学。计算机协会219 (2003)261 270。
30.A.L.佩里,P.J.低,jr埃利斯,J.D.雷诺兹,科学308 (2005)1912 1915。
31日。d·贝尔·伯恩斯,e·琼斯,k .桃子e . Portilla t·格雷格e·麦肯齐·d·里德,水珠。改变生物(2004)1209 1213。
32。k .打烙印,布罗姆·g·博尔赫斯第一,k . Erzini g·亨德森,开国元勋之一B.R.安贝德卡对麦肯齐,h·门德斯,j·里贝罗A.M.P.桑托斯r . Toresen冰3月科学。计算机协会219 (2003)261 270。
33。v.j.n Wearmouth,西姆斯M.J.创说D.W. E.J.索思豪尔,A.J.向南,P.A.亨德森,中华民国霍金斯Proc。r . Soc。Lond。爵士。B:生物。科学,271 (2004)655 661。
34。G.C.上升,冰R.L.奥德利j . 3月Sci, 59 (2002) 1018 1026。
35。J.M. Grebmeier J.E.陆路,S.E.摩尔,那么Farley。卡马克,l·库珀动向Frey J.H.赫勒,碰头麦克劳克林,S.L.麦克纳特科学311 (2006)1461 1464。
36。C.D. Harvell, k金、伯克霍尔德J.M.水银血压计Colwell,公关爱泼斯坦,D.J. Grimes一些霍夫曼,E.K. Lipp, a . Osterhaus . Overstreet, J.W.波特G.W.史密斯,广义相对论Vasta,科学285(增刊。5433)(1999)1505 1510。
37岁。d·h·McCallum Harvell, a·多布森生态。列托人。6(1062年12月,2003)1067。
38。哈里斯议员,d·贝尔,r . Toresen l . Nottestad m . Kloppmann h .杜诺k .桃子D.R.A.拉什顿,j·福斯特史密斯,s .万利斯3月杂志。151 (2007)973 983。
39岁。N.K. Dulvy S.I.罗杰斯,s·詹宁斯诉·S.R.染料,H.R. Skjodal, j .达成。生态。(2008)doi: 10.雷竞技csgo1111 / j。1365年2664.2008.01488.x
40。大梁,k . Lengfellner水珠。改变生物学观点》14 (2008)1199 1208。
41岁。g . Beaugrand K.M.打烙印,j·林德利s Souissi pc里德,自然426 (2003)661 664。
42。J.C. Molinero f . Ibanez说,s . Souissi m . Chifflet p .雪环境科学145 (2005)640 649。
43。D.L. Mackas, r·戈德布拉特A.G.刘易斯。j .鱼。Aquat。Sci, 55 (1998) 1878 1893。
44岁。千叶,M.N. Aita, k . Tadokoro t . Saino h . Sugisaki k .醒来时,掠夺。Oceanogr。77 (2008) 112 126。
45岁。g . Beaugrand pc里德,水珠。改变杂志。9 (2003)801 817。
46岁。爱德华·m·g . Beaugrand pc里德和劳登,M.B.琼斯,3月生态。掠夺。Ser。239 (2002) 10。
47岁。m·爱德华兹·里德b Planque冰j . 3月Sci。58 (2001) 39 49。
48。J.M. Fromentin, b . Planque 3月生态。掠夺。Ser。134 (1996) 1雷竞技csgo11 118。
49。水银血压计Kirby g . Beaugrand j·林德利A.J.理查森,m·爱德华兹,pc里德,3月生态。掠夺。爵士,330 (2007)31 38。
50。C.P. Lynam,中华民国干草,主管Limnol。Oceanogr。49 (2004)637 643。
51。pc里德·m·爱德华兹,Senck。玛莉特•31 (2001)107 115。
52岁。pc里德·m·爱德华兹,h·g·亨特,A.J.华纳,自然391 (1998)546。
53岁。A.J.理查森,科学博士斯科曼,科学305 (2004)1609 1612。
54。m·爱德华兹D.G.约翰,南卡罗来纳州Leterme e·斯文森主持A.J.理查森,Limnol。Oceanogr。51 (2006)820 829。
55。m . Frederiksen m·爱德华兹A.J.理查森,北卡罗来纳州韩礼德,s .万利斯j .似的。生态,1259 1268。
56。m·爱德华兹,p . Licandro D.G.约翰,A.W.G.约翰,D.P.史蒂文斯SAHFOS技术。众议员3 (2006)1 8。
57。fw整流罩,c .莫尔曼得出Hinrichsen, k维兰德,j . Tomkiewicz r·沃斯克劳斯·a . Makarchouk开国元勋之一B.R.安贝德卡对麦肯齐,硕士圣约翰,d . Schnack n . Rohlf t . Linkowski J.E.拜尔,冰j . 3月Sci 62 (2005) 1408 1425。
58岁。j . Alheit c .莫尔曼j . Dutz g . Kornilovs p .卢安克诉Mohrholz n . Wasmund冰j . 3月Sci 62 (2005) 1205 1215。
59。g . Beaugrand掠夺。Oceanogr 60 (2004) 245 262。
60。J.C. Molinero, f . Ibanez说,p .积雪,e . Buecher s Souissi Limnol。Oceanogr。50 (2005) 1213 1220。
61年。D.G.约翰·m·爱德华兹,a·理查森系统Spicer, 3月生态。掠夺。Ser。265 (2003) 283 287。
62年。T.J.史密斯,t . Tyrrell Tarrant,地球物理学。卷,31(2004)货号。L11302。
63年。莫莱森Verheye,误判率。j . 3月Sci. / S。误判率。Tydskr。Seewet 19 (1998) 317 332。
64年。B.E. Lavaniegos。欧曼、深海研究》(II。钉。Oceanogr) 50 (2003) 2473 2498。
65年。a·阿特金森诉Siegel, e . Pakhomov p . Rothery自然432 (2004)100 103。
66年。c . Barbraud h . Weimerskirch自然411 (2001)183 186。
67年。h . Weimerskirch p . Inchausti c . Guinet c . Barbraud Antarct。Sci, 15 (2003) 249 256。
68年。查尔斯C.D. D.B. cooper,顶替Baumgarter,诉费雷拉Bartrina,医学博士欧曼、科学311 (2006)63 66。
69年。千叶,k . Tadokoro h . Sugisaki t . Saino水珠。改变杂志。12 (2006)907 920。
70年。千叶,t . Saino掠夺。Oceanogr 57 (2003) 317 339。
71年。北卡罗来纳州Stenseth, a . Mysterud g . Ottersen J.W. Hurrell, m·利马K.S. Chan科学297 (2002)1292 1296。
72年。F.P.查韦斯,j . Ryan S.E. Lluch•科m . Niquen科学299 (2003)217 221。
73年。h谢长廷、C.S. Reiss r。休伊特,g .苏吉哈拉。j .鱼。Aquat。科学,65 (2008)947 961。
74年。g . Beaugrand f . Ibanez说,3月生态。掠夺。Ser。232 (2002) 197 211。
75年。g . Beaugrand f . Ibanez说j·林德利3月生态。掠夺。Ser。219 (2001) 189 203。
76年。g . Beaugrand f . Ibanez说j·林德利pc里德,3月生态。掠夺。Ser。232 (2002) 179 195。
77年。卢瑟福,s D 'Hondt w . Prell自然400 (1999)749 753。
78年。pc里德,D.G.约翰·m·爱德华兹,m·斯塔尔m .的真心拥护和诚挚谢意,p . Snoeijs水珠。改变生物13 (2007)1910 1921。
80年。D.G.卡彭,J.A.烧伤,摩根大通(J.P.蒙托亚,a, c . Mahaffey t·甘德森自动跟踪Michaels, E.J.木匠,水珠。Biogeochem。周期19 (2005)1 17。
81年。l . Bopp o·鲍彻,o . Aumont s Belviso J.L.杜福瑞斯m . Pham, p . Monfray即可。j .鱼。Aquat。科学,61 (2004)826 835。
82年。S.L.斯特罗姆,科学320 (2008)1043 1045。
继续阅读:珊瑚礁生态系统的变化,气候和全球变化的一个指标raybet雷竞技最新
这篇文章有用吗?
读者的问题
-
亚丁湾3个月前
- 回复