北极光限制
U]00动物园MM«0 500 600 TOO«00 900 llM) 1100 l!设备
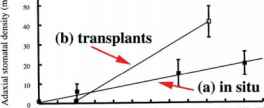
图3.9 V. myrtillus气孔密度随海拔的变化。(a)现场观测。(b) 200-900米的移植(经Woodward et al., 2002年许可转载)。
U]00动物园MM«0 500 600 TOO«00 900 llM) 1100 l!设备
高度
图3.9 V. myrtillus气孔密度随海拔的变化。(a)现场观测。(b) 200-900米的移植(经Woodward et al., 2002年许可转载)。
桃金娘的正面气孔密度随着生长的增加而增加增加高度(无花果。3.8 - -3.9)。当单株从200米移栽到905米时,有更明显的反应,显示出对海拔升高的影响有表型反应的能力。同样地,生长在雪斑的植株平均正面气孔密度为18 mm~2,而生长在裸露地区的植株平均正面气孔密度为55 mm~2。成熟叶片似乎能感知到改变叶片发育与气孔密度相关的信号,进而影响发育芽叶片中的气孔密度(Woodward et al., 2002)。
资源搜寻是功能调整的另一个方面,这体现在一些植物主动寻找资源的能力上。这在匍匐茎植物中最为明显,在这些植物中,表层茎或地下匍匐茎可以将新芽放置在有利于光线捕捉的位置,而地下匍匐茎可以帮助根系寻找矿物质营养物质。紫色石肺草(Saxifraga oppositifolia)有两种形态(章节2.2.4和3.2.4),其中一种有簇状芽和主根,不觅食,生活在干燥的栖息地,生长季节较长。另一种形式采用爬行的习性,这使它能够寻找资源,利用岩石裂缝和其他地方的庇护所,营养和水,并且在呼吸和光合作用方面有更大的代谢活动,这弥补了与潮湿的洞穴有关的较短的生长季节,那里的雪覆盖到生长季节的后期。常见的沼泽棉莎草是两个共享相邻栖息地的密切相关物种的一个例子,其中一个,Eriophorum angustifolium,是一个匍匐茎的觅食物种,而E. vaginatum是一个形成獠牙的非觅食物种。后者更能耐受地下水位深度的变化,而前者则在植物通过其耐洪匍匐茎传播的湿润地区更成功(图3.10-3.11)。
有机体越大,它所能支配的不利于竞争对手的资源份额就越大,而且往往不成比例地大。因此,大树从它们所占据的地形上吸收了大量的光线。在科学术语中,对光的竞争是大小不对称的,因为较大的植物可能在竞争关系中占据主导地位,因此较高的植物获得的光资源与其大小不成比例(Blair, 2001)。
在边际的栖息地环境往往是不确定的,干旱、洪水、风暴或寒冷的发作,加上物理干扰,可以破坏大型植物主宰景观的趋势。在这些情况下,至少在地面上的大小可能与长期生存不相容,接触较少和对资源需求较低的生物可能会占上风。因此,重要的是要考虑植物存在的双重性质,一部分在空气中,另一部分扎根于地面或土壤中
-
 colonizing a bog from which peat has been recently extracted industrially in Caithness, Scotland.")
- 图3.10在苏格兰的Caithness,沼泽棉(Eriophorum vaginatum和E. angustifolium)殖民于泥炭最近被工业提取的沼泽中。
 colonizing a bog from which peat has been recently extracted industrially in Caithness, Scotland.")
-
 The tufted non-foraging species E. vaginatum which favours drier portions of the bog. (Right) The foraging species E. angustifolium which grows in the wetter parts of the bog.")
- 图3.11沼泽棉花生长的不同策略。(左)簇状的非觅食物种,喜欢沼泽干燥的部分。(右)生长在沼泽较潮湿地区的觅食物种E. angustifolium。
 The tufted non-foraging species E. vaginatum which favours drier portions of the bog. (Right) The foraging species E. angustifolium which grows in the wetter parts of the bog.")
淹没在水中。与空中拍摄相比,植物的地下部分生活在一个潜在的更不均匀的栖息地,资源很少均匀地分散。因此,从土壤中获取资源可能更加大小对称,因为获得的土壤养分的数量将与觅食器官的大小成正比。不幸的是,大多数研究使用的是均匀分布的养分资源,而土壤均一性在自然界中并不常见。
3.2.3能力适应的不利方面
每一种对特定环境的适应都有其局限性,这些局限性来自于对特定环境条件的依赖
-
 Scot's lovage (Ligusticum scoticum). (Right) The oyster plant (Mertensia maritima). (Reproduced with permission from Hulten & Fries, 1986).")
- 图3.12在不列颠群岛和斯堪的那维亚达到其南部分布极限的两个北极海岸物种的欧洲分布。(左)Scot's lovage (Ligusticum scoticum)。(右)牡蛎植物(Mertensia maritima)。(经Hulten & Fries授权转载,1986年)。
 Scot's lovage (Ligusticum scoticum). (Right) The oyster plant (Mertensia maritima). (Reproduced with permission from Hulten & Fries, 1986).")
在适应的植物中创造。从这个意义上说,“适应是走向灭绝的第一步”(Crawford, 1989)。因此,寒冷气候的植物利用适应能力来弥补低温的限制,可以使这种适应能力强的植物在温暖的环境中处于不利地位。raybet雷竞技最新这方面的例子是一些北极物种由于温暖环境而出现的碳缺乏。北极沿海草本植物Scot's lovage (Ligusticum scoticum)和牡蛎植物(Mertensia maritima)的南部分布就是这种情况的例子。这两个物种都出现在北极(图3.12-3.15),并在苏格兰海岸达到其分布的南部极限。它们还以冬季后重新生长时扩展叶子的速度而著称。如此快速的生长,利用之前获得的碳储备,需要很高的呼吸频率。scoticum对温度呼吸速率的典型响应如图3.14所示。在牡蛎植物中也发现了暗呼吸速率随温度的快速增加,与较南部分布的两个类似沿海物种的呼吸速率较低形成对比。 The depletion of carbohydrate reserves by high respiratory activity at warm temperatures has often been suggested as a limiting factor in the southward extension of northern species (Crawford & Palin, 1981).
对生长在苏格兰的各种Vaccinium物种碳水化合物含量的研究(Bannister, 1981)表明,在温暖温度下碳水化合物储备消耗最严重的物种V. uliginosum在其南部活动范围内受到的限制最大。因此,对于一些多年生草本植物和木本石南植物来说,在短暂的凉爽生长季节中,通过高呼吸速率来利用碳水化合物储备的结果,使植物在温暖的气候中处于不利地位(另见图3.12-3.15)。raybet雷竞技最新
许多研究试图确定木本植物的总碳平衡是否可以
-
 Scot's lovage (Ligusticum scoticum). (Right) The oyster plant (Mertensia maritima).")
- 图3.13主要分布在北极和亚北极的沿海物种生长在奥克尼的一个瓦滩上。(左)Scot's lovage (Ligusticum scoticum)。(右)牡蛎植物(Mertensia maritima)。
 Scot's lovage (Ligusticum scoticum). (Right) The oyster plant (Mertensia maritima).")
相反,即树带界线附近的树木生长不受碳供应的限制(图3.16),更有可能的是,在当前环境二氧化碳浓度下,碳汇活动及其由环境的直接控制限制了树木的生物量生产(Hoch & Körner, 2005)。
虽然在木本植物总生物量中测量的碳限制可能不是与木本物种随温度分布直接相关的特征,但在特定条件下,某些重要器官,如根尖和芽以及形成层组织,而不是整个植物可能严重缺乏碳(第3.2.4节)。
3.2.4气候变暖与特定组织的脆弱性
根的分生区总是无氧的。因此,任何对根系氧气供应的减少都会增加根系缺氧的数量,从而通过无氧呼吸加速碳水化合物水平的下降。碳水化合物的转运似乎不能补充根的远端区域在仲冬。这些组织碳水化合物的消耗降低了抗氧化剂的含量,从而使根系在地下水位下降和恢复通气时容易受到缺氧后损伤
继续阅读:信息之杰
这篇文章有用吗?