念珠藻属的苔藓植物共生的实验系统
本章将审查结果的研究主要是利用一个协会之间的固氮藻青菌念珠藻属punctiforme(写明ATCC 29133) 73102年应变PCC,同义词和苔藓植物金鱼藻Anthoceros毛虫,经常在实验室重组与单独pure-cultured伙伴。将更加关注机制的相互作用比共生体的多样性;后者是一个扩大的话题最近研究使用分子遗传技术(西部和亚当斯1997;科斯塔et al . 2001;拉斯穆森和尼尔森2002)。的完整基因组序列n punctiforme写明ATCC 29133目前已知(应变http://genome.jgi-psf.org/microbial/)。因此,将讨论结果和概念和假设提供n . punctiforme基因组序列的上下文以及遗传、生理生化特征的两个合作伙伴。在适当的时候,比较蓝藻也将对其他的-植物系统。
念珠藻属punctiforme有着非常广泛的生理属性,营养细胞发展替代能源和生态位(米克斯et al . 2002;米克斯2003)。适合基因操作,包括随机转座子突变(科恩et al . 1994年)和有针对性的基因替换(哈根和米克斯1999)。这些共同特征提高其基因组序列的科学价值(米克斯et al . 2001年)。n punctiforme代表一个组在完全黑暗的蓝藻生长呼吸异养生物,代谢模式,可能需要在共生协会。发展替代的两个至关重要的共生互动;丝的分化等hormogonia,作为植物协会的感染性单位,和异形胞的,唯一的固氮酶表达在几乎所有念珠藻属菌株(图1)。第三个营养细胞发育替代是静息细胞称为厚壁孢子的形成。厚壁孢子可以检测到开始衰老地区植物协会,但他们的角色,如果有的话,在共生互动尚未解决。包括营养细胞周期的四个发展方向n punctiforme营养细胞中数值无与伦比的细菌。他们提供了多个阶段发育的细胞周期调控,包括环境共生伙伴的信号。n . punctiforme基因组包含一个非常大量的感官转导系统与大约156传感器组氨酸激酶,103响应调节蛋白质,和51个丝氨酸/苏氨酸蛋白激酶,+ 7的腺苷酸/鸟苷酸环化酶(米克斯et al . 2001;米克斯2005)。环境信号,调节这些感官系统的活动在很大程度上是未知的;然而,有足够的调节蛋白作为潜在的目标信号的共生伙伴。
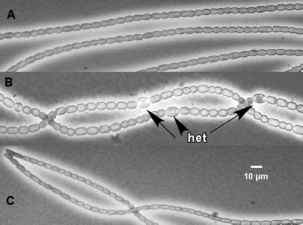
图1所示。相衬显微照片的三个n . punctiforme田鼠自然发展状态。一铵丝没有任何分化细胞增长。B Di-nitrogen成长纤维含有固氮异形胞(het)出现在一个间隔的非随机模式。C能动的连锁体丝显示小细胞大小相对于氮气或NH4 +种植文化。板A和C复制从米克斯et al . (2002)
图1所示。相衬显微照片的三个n . punctiforme田鼠自然发展状态。一铵丝没有任何分化细胞增长。B Di-nitrogen成长纤维含有固氮异形胞(het)出现在一个间隔的非随机模式。C运动型连锁体细丝显示小细胞大小相对于氮气或NH4 +种植文化。板A和C复制从米克斯et al . (2002)
念珠藻属punctiforme显示广泛的共生在植物的系统发育谱能力,包括苔藓植物角苔纲(Enderlin和米克斯1983)和苔类(约瑟夫和亚当斯2000),裸子植物苏铁植物,从那里这是孤立(Rippka et al . 1979年),和被子植物Gunnera spp。伯格曼(约翰逊和1994),以及non-lichen真菌,g . pyriforme(克鲁格et al . 2002年)。植物协会的照片,说明不同的隔间藻青菌的本地化,图2中给出。很难想象,n . punctiforme先后开发了具体的自适应过程对于每个植物组的成员组出现在进化的时间。相反,它是合理的假设,植物必须有独立进化而来的策略来控制键监管和藻青菌的代谢途径,通常表现在独立生存的增长。例如,转向一个异养的碳和能量代谢方式,hormogonia和异形胞的分化N2固定念珠藻属的基本过程表达除了植物伙伴,但反应增强的工厂合作伙伴(米克斯1998)。因此,从独立生存的过渡到共生念珠藻属的增长可以被最好地概括为响应度的变化。相比之下,根瘤菌与豆科植物时,植物细胞变成一个类细菌状态和诱导固氮酶合成和活动,无论是他们的表达除了植物伙伴(van Rhijn和Vanderleyden
1995)。这些形态和生理特征转换rhizobial共生增长状态最好的一种新型反应。此外,念珠藻属殖民结构或区域(称为共生蛀牙)植物的存在和变化很少在共生状态。相反,根瘤菌诱导根瘤的形成,它可以被视为一个新的植物器官、结构上防止固氮酶氧气,同时提供足够的氧气rhizobial呼吸(van Rhijn和Vanderleyden 1995)。念珠藻属,和相关heterocyst-forming物种,携带自己的氧气保护机制可能占相对简单的共生关系。程度上的这些差异,而响应,和简单的结构,和一个新的器官,导致我们的结论Nostoc-based的互动共生,主要(但不仅限于)单向从植物到藻青菌(米克斯1998),与合作伙伴之间的广泛的信号交换特征rhizobia-legume协会(Perret et al . 2000年)。
裸子植物苏铁属植物苏铁
裸子植物苏铁属植物苏铁
-

- 苔藓植物的homwort Anthoceros的被子植物Gunnera


图2所示。的照片代表的三个地面组植物伙伴n punctiforme将形成共生关系。裸子植物苏铁sp.显示植物的叶子(A)和(B)一个切除的珊瑚状的根叶分段显示蓝藻环形腔(nc)。酒吧板A和B是0.5和0.5厘米,分别。苔藓植物的金鱼藻Antho-ceros毛虫显示配子体(G)和附加孢子体C和D (S)代面板在实验室纯培养的配子体组织重建与n punctiforme局部粘液蛀牙(nc)。板C和D的酒吧都是1.0厘米。被子植物的Gunnera种虫害描绘与独特的幼苗茎腺体(E)念珠藻属的网站进入阀杆和(F)干的一个巨大的弦切面Gunnera chilensis植物展示许多共生蛀牙被念珠藻属(nc)。面板E和F的酒吧都是1.0厘米。从米克斯和复制Elhai (2002)
图2所示。的照片代表的三个地面组植物伙伴n punctiforme将形成共生关系。裸子植物苏铁sp.显示植物的叶子(A)和(B)一个切除的珊瑚状的根叶分段显示蓝藻环形腔(nc)。酒吧板A和B是0.5和0.5厘米,分别。苔藓植物的金鱼藻Antho-ceros毛虫显示配子体(G)和附加孢子体C和D (S)代面板在实验室纯培养的配子体组织重建与n punctiforme局部粘液蛀牙(nc)。板C和D的酒吧都是1.0厘米。被子植物的Gunnera种虫害描绘与独特的幼苗茎腺体(E)念珠藻属的网站进入阀杆和(F)干的一个巨大的弦切面Gunnera chilensis植物展示许多共生蛀牙被念珠藻属(nc)。面板E和F的酒吧都是1.0厘米。从米克斯和复制Elhai (2002)
deBary最初定义的(1879),这个词共生简单地描述了生活在一起的不同命名的有机体。现在最常应用于互惠互动中涉及的所有生物明显受益于亲密的物理协会,与寄生交互中一方受益,另一个是慢慢地伤害。互惠的术语可能适当地反映异养根瘤菌的rhizobia-legume协会受益于一个物理协会能光合自养的植物,而植物得益于固氮细菌的活动。的好处能光合自养的念珠藻属与能光合自养的工厂是不明显,除了,也许,作为庇护以避免放牧捕食。作为替代互惠和寄生虫,我们已经描述了共生固氮念珠藻属协会作为一种更准确的概念化的植物驯化蓝藻ammonium-producing工厂(共生体)(米克斯和Elhai 2002)。藻青菌没有好处也没有受到伤害的协会。因此,我们正在测试的工作模型并不吸引监管或结构基因类比rhizobial有节(点头)基因活动只在协会的细菌与豆类(van Rhijn和Vanderleyden 1995;Perret et al . 2000),尽管通用策略和化学信号可能是相似的。
上面的信息单向流动的概念化共生的相互作用会导致监管电路和结构基因的假设目标在这些固氮蓝藻协会唯一进化的血统。我们提出一个类似的进化场景上下文中的营养发展替代能源(米克斯et al . 2002;米克斯2005)。因此,基因编码的过程不能由纯粹的比较基因组序列中确定生物信息学方法。相反,功能分析将鱼各自所需基因的基因组。
作为指导在设计功能的分析,我们已经开发出工作模型。n . punctiforme之间的交互和a .毛虫,而最有可能的操作作为一个连续体,可以实验分为两个连续的阶段,每个子阶段(图3)。协会和涉及到的初始阶段是建立连锁体细丝的分化和行为感染单位。第二阶段是开发的功能涉及到植物细胞的生长和代谢改变,和异形胞的分化和行为。这个模型是我们支持的突变体的n . punctiforme孤立单独建立或功能性的缺陷发展协会(表1)。突变基因的生理角色的产品将在随后的章节中讨论。
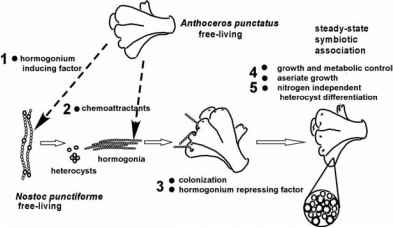
图3所示。原理之间的相互作用的连续a蜥和n puncti-forme导致N2-fixing共生协会。的交互是藻青菌描绘成单向从植物。数字指文本中所描述的实验不同的子阶段。复制修改与米克斯(2003)
图3所示。原理之间的相互作用的连续a蜥和n puncti-forme导致N2-fixing共生协会。的交互是藻青菌描绘成单向从植物。数字指文本中所描述的实验不同的子阶段。复制修改与米克斯(2003)
应变 |
共生 |
乙炔 |
基因 |
阶段 |
|
coloniesa |
reductionb |
诱导 |
影响 |
||
byc |
|||||
每g弗兰克-威廉姆斯 |
殖民地 |
||||
写明ATCC 29133 d |
0.21±0.04 |
6.3±1.2 |
12.4±3.3 |
- - - |
- - - |
UCD 398(叹息)d |
1.2±0.2 |
8.0±3.9 |
10.1±4.1 |
低氧诱导因子 |
感染 |
UCD 328 (hrmAf |
1.6±0.1 |
6.1±1.1 |
8.6±1.3 |
HRF |
感染 |
UCD 444 (ntcA) f |
0 |
0 |
0 |
- - - |
感染 |
UCD 416 (hetF) |
0.26±0.06 |
0 |
- - - |
函数 |
|
UCD 464 (hetR) f |
0.36±0.04 |
0 |
0 |
- - - |
函数 |
“共生的殖民地在解剖显微镜可见培养2周后,规范化毫克干wt a毛虫的组织统计和| g的背影的念珠藻属变质剂
本现场乙炔还原活性作为nmol乙烯形成每克鲜重配子体组织或每共生pmol乙烯形成念珠藻属殖民地
cHIF是连锁体诱导因素,指渗出物a毛虫的包含因子(s),引发连锁体分化;HRF连锁体抑制因素,指的是一种水提物a毛虫的包含因子(s),压制连锁体分化”坎贝尔et al。(1998) eCohen和米克斯(1996)fWong和米克斯(2002)
“共生的殖民地在解剖显微镜可见培养2周后,规范化毫克干wt a毛虫的组织统计和| g的背影的念珠藻属变质剂
本现场乙炔还原活性作为nmol乙烯形成每克鲜重配子体组织或每共生pmol乙烯形成念珠藻属殖民地
cHIF是连锁体诱导因素,指渗出物a毛虫的包含因子(s),引发连锁体分化;HRF连锁体抑制因素,指的是一种水提物a毛虫的包含因子(s),压制连锁体分化”坎贝尔et al。(1998) eCohen和米克斯(1996)fWong和米克斯(2002)
继续阅读:协会成立Hormogonia分化和行为
这篇文章有用吗?
读者的问题
-
Aziza1年前
- 回复