丙氨酸合成和分泌的根瘤菌Leguminosarum
微生物学,AMS学院读大学Whiteknights,阅读,RG6 6 aj,英国
15 n-labeling研究的基础上,它被普遍接受,铵是唯一的分泌的产物N2-fixation类细菌和植物负责同化成氨基酸。然而,15 n-labeling研究表明大豆类细菌分泌丙氨酸和氨(水域et al . 1998年)。我们调查这个豌豆类细菌,发现铵和丙氨酸都是分泌。
丙氨酸分泌可能占到20%的丙氨酸的分泌和铵(Allaway et al . 2000年)。它们之间的体外分区将取决于系统是否打开或关闭,以及铵浓度和类细菌密度。这可能是解释为铵的低亲和力的丙氨酸脱氢酶,被确定为5.1毫米。类似于整个细胞排泄丙氨酸与铵的公里3.2毫米,因此体外检测丙氨酸合成的能力取决于试验系统中铵的积累。
丙氨酸脱氢酶的活性较高的大豆和豌豆类细菌相比,可能有利于丙氨酸合成大豆。然而,由于实验条件可以改变丙氨酸由孤立的类细菌的数量我们识别和丙氨酸脱氢酶的基因突变(艾达)。这证实,艾达是在孤立的类细菌丙氨酸合成的主要途径。类细菌的艾达突变修复只是分泌铵氮的比率相当高,导致降低总氮分泌。豌豆接种aldA突变是绿色和健康证明铵类细菌分泌的可以为植物生长提供足够的氮。植物接种的变异减少生物量与野生型相比,接种。标签和植物生长的研究表明,丙氨酸合成和分泌有助于N2-fixation因此生物质积累的效率。
我们目前过度表达AldA测试假设的艾达将是至关重要的丙氨酸类细菌形成的。完整的表达AldA似乎要求aldR和艾达都是出现在一个质粒,大概由于aldR自动调整(见下文)。
发散思维从艾达是一个亮氨酸转录监管机构蛋白质(单体),同系物aldR。插入的interposon aldR阻止转录的艾达,以gusA融合和Northern,以及艾达酶活性。AldR是自动调整的,因为AldR插入也防止aldRr转录。gusA融合。艾达活动引起的增长对羧酸,包括丙酮酸、苹果酸和琥珀酸和丙氨酸。我们正在调查丙氨酸是否由独立生存的文化和如何排出氧气张力等因素可能影响这个和增长阶段。丙氨酸似乎没有分泌独立生存的文化在指数期,但一阵丙氨酸合成可以在固定相感应L-malate过剩和铵的文化。
丙氨酸合成也可以被视为一个可能的溢出途径在同样意义上的合成polyhydroxybutyrate (PHB)和糖原。我们因此被调查变异的影响每个通道单独在一起。这也是做的不确定和确定植物(豌豆和豆类)使用同基因的父母只有符号不同菌株质粒。糖原合酶突变体(glgA)显示大量增加植物淀粉的结节但仍能固氮。这表明,糖原合成在豌豆类细菌可能是一个主要的碳存储化合物能够深刻影响碳代谢在植物。突变体的两倍和三倍的建设应该帮助我们了解这些途径是塑料。我们特别想知道是否阻塞一个或多个通路引起的碳和转移还原剂其他溢出途径。
引用
Allaway D et al。(2000)摩尔。Microbiol。36岁,508 - 515
水域JK et al。(1998) Proc。国家的。学会科学。美国95年,12038 - 12042
同化的FIXED-N UREIDE-FORMING共生关系
西澳大利亚大学的植物学部门,Crawley佤邦6009年,澳大利亚
马2部门的生物学、麻省理工学院、剑桥大学,02139年,美国
最近的数据产品的长期持有的观点提出了挑战固氮酶活动、氨、转移从symbiosome到宿主细胞胞质,同化(了天et al . 2001年)。水域et al。(1998)提出了证据固定的排泄N为丙氨酸类细菌分离大豆(大豆[L。]稳定)通过蔗糖密度梯度分离和纯化。然而,排泄和积累的丙氨酸类细菌在p02小于0.01,最高0.06 p02的固氮酶活性最大。另一方面,flow-chamber固氮酶活性优化的实验条件下,也有隔离类细菌从大豆,认为氨的主要排泄产品固定(李等人。2001)。显著区别这两种类型的实验方法是切除排泄时产品主要材料的固定系统积累的封闭系统所使用的水域et al .(1998)和Allaway et al。(2002)表明,这是解释相互矛盾的结果。
-
")
- 30 40 50时间(分钟)
")
图1所示。固定的N2化验15 N2标签的结节状的根豇豆。数据意味着±SE (n = 5)和线由non-lmear回归拟合。大气中m每个孵化船化验同位素浓缩前收集根系,发现在% 74.7±0.5 x 15 n (n = 30)。
目前的研究报告的结果稳态I5N2标签完好无损的结节状的根豇豆植物(豇豆属unguiculata (L。Walp。简历个人简历3:Bradyrhizobium应变CB756)的情况下,保持高水平的固氮酶活性(250 p。摩尔N2 h“1 g”1 DW结节;图1)。根系被冻结在液体N2 5、10、20、30、45或60分钟,结节切除。五次重复了同样的实验,并提取溶质用气相色谱-质谱的分析同位素浓缩TBDMS衍生品。图2显示快速标签的酰胺基谷氨酰胺(single-labeled分子)达到平衡在ca。50%的同位素浓缩后15 n2提供5分钟。Double-labeled分子(酰胺和氨基酸组)积累较为缓慢,符合标签的传输较慢,谷氨酰胺的氨基。在豇豆结节,如大豆,使用谷氨酰胺的酰胺基的两个amidotransferases新创嘌呤通路形成IMP对黄嘌呤氧化和酰(史密斯对阿特金斯,2000)。double-labeled黄嘌呤分子(图3)的积累速率大于single-labeled分子或3或4 15 n原子与这个途径是一致的。此外,稍微慢标签动力学的黄嘌呤预计将中间的通道16酶步骤从第二glutamine-dependant步骤(FGAR amidotransferase)。4个氨基酸的标签如图4所示是一致的与他们收购的氨基N组通过GOGAT谷氨酰胺转氨酶的活性。
尽管这项研究无法考虑的影响,这些化合物可能的多个池标签动力学显示没有证据支持这个想法,丙氨酸是一种前体的谷氨酰胺的酰胺基或嘌呤环。
40-j |
||||
■o |
||||
V |
•问 |
30 - |
||
类风湿性关节炎 |
z |
3 |
在 |
|
£ |
20 - |
|||
03 |
年代 |
|||
O |
||||
b |
10 - |
|||
年代? |
U - 1 - |
||
一个米 |
glutamme-l 15 n |
|
1 glutamine-2 15 n |
||
/我 |
||
图2。15 n标记的谷氨酰胺豇豆在接触15 n2的结节。数据意味着±SE (n = 5)和非线性回归的线路安装。
图3。15 n标记的黄嘌呤在结节豇豆J接触15 n2。数据E意味着±SE (n = 5)和^线拟合非线性=£回归。年代
图3。15 n标记的黄嘌呤在结节豇豆J接触15 n2。数据E意味着±SE (n = 5)和^线拟合非线性=£回归。年代
-
")
- 时间(分钟)
")
麸胺酸天冬氨酸谷氨酸glycire天冬氨酸glycire
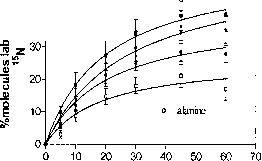
图4。15 n标记的氨基酸豇豆在接触15 n2的结节。数据意味着±SE (n = 5)和非线性回归的线路安装。
时间(rrin)
图4。15 n标记的氨基酸豇豆在接触15 n2的结节。数据意味着±SE (n = 5)和非线性回归的线路安装。
时间(rrin)
继续阅读:nif操纵子的组织
这篇文章有用吗?