树木生长在森林中
大多数研究的发展热带森林的树木发现,绝大多数的树木生长非常缓慢。例如,64%的树木在情节Bukit Lagong和河Menyala在马来西亚半岛增长率平均直径约1毫米年20-30-year ~(图3.3)
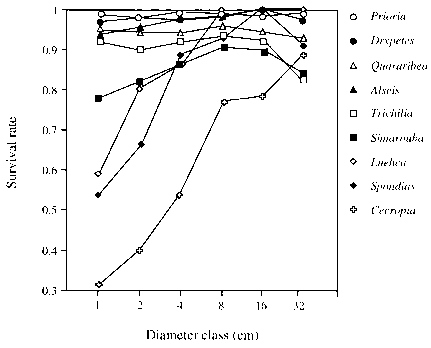
图3.2部分的植物生存intercensus间隔198285样品直径的函数类树种Barro科罗拉多岛的阴谋。许多那些在物种显示高和相对稳定的生存在所有大小类最大的除外。这类物种的例子有Prioria copaifera, Drypetes standleyi, QQuararibea aster-olepis, Alseis blackiana和Trichilia构叶。相比之下,shade-intolerant物种如Luehea seemannii, Spondias mombin和Cecropia旧址都显示增加生存随着直径的增加,接近的高存活率林分物种一旦shade-intolerant物种到达标签树冠的环境。在哈贝尔和福斯特(1990)。
图3.2部分的植物生存intercensus间隔198285样品直径的函数类树种Barro科罗拉多岛的阴谋。许多林分物种显示高和相对稳定的生存在所有大小类最大的除外。这类物种的例子有Prioria copaifera, Drypetes standleyi, QQuararibea aster-olepis, Alseis blackiana和Trichilia构叶。相比之下,shade-intolerant物种如Luehea seemannii, Spondias mombin和Cecropia旧址都显示增加生存随着直径的增加,接近的高存活率林分物种一旦shade-intolerant物种到达标签树冠的环境。在哈贝尔和福斯特(1990)。
-
 in growth rate classes in three primary dipterocarp forests in Peninsular Malaysia. After Manokaran & Kochummen (1994).")
- 图3.3频率的树(> 10 cmdbh)增长率类在马来西亚半岛的三个主要的龙脑香科树森林。后Manokaran & Kochummen (1994)。
 in growth rate classes in three primary dipterocarp forests in Peninsular Malaysia. After Manokaran & Kochummen (1994).")
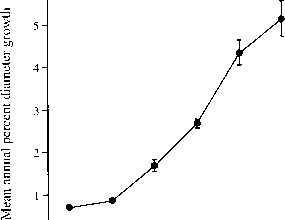
测量时间(Manokaran & Kochummen 1994)。同样,树苗1-4cmdbh Barro科罗拉多岛上显示增长率的负指数分布频率为三年的数据(Welden et al . 1991年)。的模态类是零增长。总之,大部分雨林的树木都几乎没有增长,甚至有些萎缩。被掉落的碎片击中,折断的大型哺乳动物或受到害虫攻击或疾病都可以导致领导人拍损失和减少大小,小树是克拉克(Clark & 1991)最敏感。大树一般的增长速度(图3.4),即使增长表示相对于原始大小的基础上,可能是因为更大的个人通常更明亮的条件。
个别物种的增长率在雨林相差很大,平均直径增长率普遍在0.5年~ 6毫米,最大速度达到年~ 15毫米(图3.5)。连续增量之间往往有强烈的自相关测量林木(斯et al . 1987年)。种植者仍停滞在增长缓慢,而主要树木保持高增长率。有几个可能的原因。最明显的是fast-growers占据特别有利的网站通常在森林而缓慢的不利环境的森林内部。树需要供应充足的光线,水分和养分迅速增长。土壤水分和养分有效性做展览的时间和空间分布森林地面,但不是一致的,大光供应率的变化。在昏暗的森林火情大多数树个人坐落在光线不足的快速增长。只有个人冠树冠的顶部附近,或位于缺口,可能得到足够的光来实现高的增长率。树木的深沉强烈抑制经济增长。他们可能失去反应能力如果更多的光变得可用,加强增长的时间自相关。几乎没有研究长期阴影抑制对树生理的影响。内在增长率的树木也一定程度的遗传控制和树木是快速增长的遗传易感性。识别这种树木一直是林业工作者的野心。然而,领导
类(胸径,厘米)直径
图3.4年平均增长率所有个体的百分比树冠树种在50公顷Barro科罗拉多岛上。这些意味着未加权的:总增长增量除以总数量的个体。有些植物减少直径由于阀杆断裂;这些植物被排除在外。竖线表示95%可信限。限制为1 - 2和2 - 4厘米胸径类太接近圈可见(样本大小超过000个人在这些大小类)。在康迪特等等。(1992)。
-
, Ashton & Hall (1992), Lieberman et al. (1985a) and Korning & Balslev (1994).")
- 图3.5平均直径增加对最大增量有七个热带森林树种的网站。树> 10 cmdbh n > 20。数据从Manokaran & Kochummen(1994),阿什顿和霍尔(1992),利伯曼et al。(1985)和korn & Balslev (1994)。
, Ashton & Hall (1992), Lieberman et al. (1985a) and Korning & Balslev (1994).")
树不一定是优越的。快速增长,例如,可能会降低抵抗害虫和疾病的成本。
树性能与光气候raybet雷竞技最新
不容易获得定量数据之间的关系生存或增长率和事件辐照度大热带树木由于技术上的困难获取长期的气候数据高于高大的树木。一种方法用来绕过这是采用半定量或定性的措施的相对光可用性树木和使用这些比较个人的增长率。不同冠照明指标可用克拉克(Clark & 1992)。确立在视觉评估这些涉及一个或多个接收到的光量皇冠每棵树的研究在预定的范围内,从“皇冠完全清楚的从太阳在各个方向的“皇冠连续树冠之下”。六棵大树物种的详细研究了克拉克&克拉克(1992)在热带雨林,所有显示冠照明指数的增加而增加的个人(图3.6)。物种之间的最大区别是失败的Minquartia guianensis达到高crown-illumination-index分数。这个物种不会达到这样大的尺寸为他人,因此无法长大从而获得皇冠不晦涩的四面八方。
利伯曼et al。(1995)使用的几何分析冠高度和距离焦郁闭的树木来生成一个索引。他们退化关闭指数对1000 /胸径获得线性关系,和
-
.")
- 图3.6冠照明指数连续大小类六棵大树物种在La热带雨林,哥斯达黎加。数据从克拉克&克拉克(1992)。
.")
-
.")
- 图3.7频率分布郁闭意味着剩余的104年最丰富的树种研究利伯曼et al . (1995)。
.")
然后用线性回归的残余size-compensated估计的相对照度,这可以平均每个物种的个体。频率分布的平均剩余的104个物种研究在图3.7中给出。九个物种,包括Hampea appendiculata和Cecropia obtusifolia,残差大大高于所有其他物种的总和。换句话说,他们被发现在网站显著比平均水平。小群5种占据网站比平均水平,肮脏与Faramea terryae shade-lover最极端。
在50公顷情节Barro科罗拉多岛,森林被分为高收入和low-canopy(10米)网站(总额的12.7%)在5 m x 5 m的网格(Welden et al . 1991年)。这个森林光环境非常粗略的分类已经用来生存调查阴影的影响,增长和招聘的树苗平民物种(表3.2)。raybet雷竞技最新相对较少的物种表现出显著差异之间的生存高——和low-canopy网站。增长表现出更强的效果,所有的物种,显示网站类型之间的显著差异在low-canopy网站更快的增长。招聘胸径1厘米类表现出类似的模式,只有两个物种(Drypetes standleyi和Rheedia acuminata)
表3.2。树苗生存、增长和招聘与树冠高度Barro科罗拉多岛上在此期间1982 - 85
列表的物种数量显示显著更大生存、生长或招聘low-canopy(无叶10米以上高度)网站,遮盖网站,或者没有区别网站(冷漠)。树苗是胸径1 - 4厘米,除灌木物种(成人高度4米,胸径树苗被划分为1 - 2厘米的地方。
列表的物种数量显示显著更大生存、生长或招聘low-canopy(无叶10米以上高度)网站,遮盖网站,或者没有区别网站(冷漠)。树苗是胸径1 - 4厘米,除灌木物种(成人高度4米,胸径树苗被划分为1 - 2厘米的地方。
Low-canopy网站 |
遮盖网站 |
冷漠 |
||||
不。spp。 |
% ofspp。 |
不。spp。 |
%的spp。 |
不。spp。 |
%的spp。 |
|
生存 |
6 |
4.0 |
19 |
12.8 |
123年 |
83.1 |
增长 |
66年 |
57.4 |
0 |
0 |
49 |
42.6 |
招聘 |
70年 |
44.9 |
2 |
1.3 |
84年 |
53.8 |
数据从Welden et al . 1991。
数据从Welden et al . 1991。
更好的遮盖的招聘网站。大部分的物种对树冠类型在一个或多个人口的措施。Welden et al。(1991)认为,这意味着大多数物种“gap-neutral”他们再生生态。穷人的准确性和歧视水平光气候可能从方法用于估计可能是一个准备解释“gap-positive”物种的相对罕见。raybet雷竞技最新
死亡率、增长和成人的大小
Barro科罗拉多岛上物种被分成四个身材类基于最大高度。这些团体被灌木(4 m成人身高),“treelets”(4 - 10 m),中型树(10 - 20米)和大树(20 m成人身高)。灌木的种类显示平均死亡率约两倍的树木和“treelets”为个人1 - 9.9厘米胸径(康迪特et al . 1995年)(表3.3)。灌木的倾向于占据这个尺寸范围的低端,和一般小碎片下降可能更容易死亡。此外,“灌木”的生殖活动可能耗尽他们的资源,使他们更容易死亡。树的树苗物种更有可能表现出积极的增长和招聘应对low-canopy网站比两个最小的大小类(Welden et al . 1991年)。例如,66%的树种有显著积极回应树苗成长low-canopy网站,但只有41%的‘treelet’和‘灌木物种也是这么做的。可能更小的平均大小的‘treelet’和‘灌木树苗可能意味着两者之间的差异在光气候经历了树冠高度类不太明显。raybet雷竞技最新另一个可能的解释是下层木物种繁殖的倾向而不是种植更多在缺口。
表3.3。年平均死亡率(%)跨物种不同身材在50公顷的树木Barro科罗拉多岛,巴拿马n,物种的数量。
表3.3。年平均死亡率(%)跨物种不同身材在50公顷的树木Barro科罗拉多岛,巴拿马n,物种的数量。
胸径地位类茎10 - 99毫米 |
n |
茎> 100 mm胸径 |
n |
20米的成年身高3.2±0.4 |
71年 |
1.9±0.2 |
63年 |
成人身高2.7±0.4 10 - 20米 |
54 |
3.3±0.2 |
49 |
4 - 10米的成年身高2.9±0.6 |
41 |
2.9±0.8 |
16 |
4米的成年身高6.3±0.9 |
28 |
- - - - - - |
- - - - - - |
所有3.5±0.3 |
194年 |
2.6±0.2 |
128年 |
康迪特et al .(1995)的数据。
康迪特et al .(1995)的数据。
身材类 |
Bukit Lagong |
河Menyala Pasoh |
紧急 |
1.11 |
1.44 - 1.65 |
主要的树冠 |
1.33 |
1.84 - 1.90 |
下层木 |
1.45 |
2.58 - 2.35 |
数据从Manokaran &斯(1994)。
数据从Manokaran &斯(1994)。
大树物种显示显著较高的生存10厘米茎中胸径或更高Barro科罗拉多岛上物种比规模较小的成年身高(康迪特et al . 1995年)(表3.3)。在热带雨林,六个紧急物种Clark &克拉克(1992)的研究显示低树>年度死亡率10厘米胸径(0.44%)相比,森林的平均水平(2.03%)。同样的模式在低地龙脑香科树森林的研究在马来西亚(Manokaran & Kochummen 1987)(表3.4),和树木在厄瓜多尔(korn & Balslev 1994)。紧急的物种也表现出一些最大增长率最高的森林(Manokaran & Kochummen 1994)。图3.5的数据已经re-plotted指示有关物种的地位(图3.8)。显然,大树往往有更高的平均和最大增长率,特别是当与下层木物种。Subcanopy物种往往更变量,与一些有非常快速增长。有趣的是,盖式车顶物种估计的平均利率从定期生长量数据对个人树在森林里是类似于长期平均水平(1 - 6毫米年~)计算通过使用c年代20岁的大树在亚马逊(钱伯斯et al . 1998年)。大树的最大增长率(5 - 15毫米年~)在森林里仍然达不到相似物种的利率arboreta或种植园,在增量大于10毫米直径年~是很常见的,和超过20毫米年~是已知的(例如,数据的Ng &唐1974)。
最大预期寿命估计树木从热带雨林,哥斯达黎加,利伯曼(et al . 1985年)和厄瓜多尔(korn & Balslev 1994)显示增加长寿的趋势日益提升的地位(表3.5)。只有在数据从热带雨林,身材类之间有显著差异。这里的下层木物种明显比其他两个更短的寿命。然而,所有的声望值的范围很大类,每个高度集团拥有一些快速增长的短暂的物种。
生长和死亡数据识别物种的使用群体
经常批评的功能分类的热带树种主要是基于特别的印象有关物种而不是详细的定量信息。然而,有几次对物种利用增长和死亡率数据进行分类。
50公顷的地块研究Barro科罗拉多岛是最大的数据集
-

- 图3.5,图3.8相同的情节,但这里的符号表示每个物种的地位类。

表3.5。最大预期寿命(最大直径10厘米胸径)树种的不同地位类从哥斯达黎加和厄瓜多尔
表3.5。最大预期寿命(最大直径10厘米胸径)树种的不同地位类从哥斯达黎加和厄瓜多尔
身材类 |
哥斯达黎加 |
厄瓜多尔 |
下层木 |
126±56 |
199±103 |
(52 - 221) |
(69 - 348) |
|
13 |
7 |
|
subcanopy |
242±81 |
270±175 |
(78 - 338) |
(54 - 529) |
|
11 |
10 |
|
树冠 |
206±102 |
299±40 |
(78 - 442) |
(250 - 353) |
|
21 |
5 |
数据从利伯曼et al .(1985)和korn & Balslev (1994)。
数据从利伯曼et al .(1985)和korn & Balslev (1994)。
用于这种目的(康迪特et al . 1996年)。五个参数被用来执行主成分分析(PCA)对142年的物种。五个重要的统计工作是:年度死亡率(避免干旱年)1 - 9.9 - cmdbh > 10 cmdbh类;平均增长率为1-2cmdbh和胸径大小类10 - 20厘米;和殖民指数(招聘low-canopy网站发生的比例)。下层木物种必须处理与别人分开,因为他们没有足够的数据规模更大的树。这两个结果PCAs产生类似的结果,尤其是第一轴。大部分的物种发生在一个紧结在范围1 - 1轴,散射的物种从1到6。这些物种具有高死亡率、高增长率和倾向于招募缺口。这组是任意地分为“先锋”第一维分数超过3和构建阶段的物种范围1 - 3。 The pioneers included Cecropia insignis and Zanthoxylum belizense among the large-statured species and Croton billbergianus and Palicourea guianensis from the understorey group. An important point to note is that the species showed no marked discontinuity of distribution along the耐荫轴,作者提出的分组被定义在任意时间间隔。
在热带雨林,利伯曼et al。(1985)使用他们的增长轨迹模拟技术来估计潜在的寿命最多的树种13年的枚举数据12.4公顷的森林。他们确定了四种组织的三维图最大胸径(情节)衡量,最大潜在寿命(从最慢的增长预测仿真)和最大增长率从增长最快的模拟(估计)。这些都是:
组我:下层木物种最大增长速度缓慢,寿命短。
组II:林分subcanopy物种达到两次只要下层木物种但有相似的最大增长率。
第三组:树冠和subcanopy物种阴影宽容但可能快速增长在高光照条件和长寿。
第四组:shade-intolerant物种快速增长和短寿命。
整个物种要么是相对那些mid-canopy树木或大盖式车顶的物种。之间的区别组I和II和III和IV,似乎相当武断的外部观察者。两双组之间出现的清晰。
这两项研究,实证观察其他热带生态学家,导致这样的结论:团体基于生长和存活的物种,特别是与发光机制,成熟的身材和可能的寿命可以承认在热带雨林的树社区。这些团体不是离散,但更好的视为区域引用字符可用空间中的位置。利伯曼的四组et al。(1985)似乎是合理的,可以描述如下。
下层木物种:small-statured成熟度相对较低的最大增长率和相当高的死亡率(尽管不是高达的先驱)。一般阴影宽容与有限的增长对辐照度增加。
Subcanopy物种:中间下层木和树冠物种之间的地位。通常那些在相对较低的死亡率。一些物种长寿。
树冠物种:最大的树,通常与高青少年生存和大量增加增长率高暴露。
先锋物种:快速增长,死亡率高,尤其是在树荫下和在青少年阶段。相对较短。
这些团体,和可能的细分,在第六章更详细地讨论。
物种的相对性能相似的生活历史
Clark &克拉克(1992)详细调查了六个物种的生长和死亡在150公顷的森林地区热带雨林,哥斯达黎加。每个六将放置在上面的树冠集团的计划。所有个人遇到大于50厘米高在调查中。这些监测每年对生存和增长和评估冠照明类和森林增长阶段。使用这些数据,和一些额外的观察Simarouba amara和两种Cecropia,作者确定了四种组织如下。
A组:中发现低冠照明类和高成熟阶段的青少年的比例森林[Lecythis ampla和Minquartia guianensis]。
B组:稳定增加照明类和比例在早期阶段增加大小(Dipteryx panamensis和Hy-menolobium mesoamericanum]。
C组:倾向于在低照明类中型少年,被克拉克&克拉克作为要求差距树苗,但倾向于成为溢在建设阶段,然后等待一个新的差距事件生长[Hyeronima alchorneoides和Pithecellobium线虫)。
D组:占据最亮的网站在所有大小类(Cecropia spp。)
组一个物种显示树苗死亡率和冠照明没有显著关系。生存的其他物种却表现出改进的机会为树苗生长在光明的条件。然而,在没有太大区别的最大增长率6驾驶室天窗/紧急物种。这个问题导致Clark &克拉克认为,之间有一个权衡能力持续在树荫下,最大增长率高的光。我们将在后面的小节中回到这个主题。一般来说,生长和存活有相对小的不同反应物种之间的森林环境,特别是五紧急。在研究Welden et al .(1991),这可能是由于方法论上的问题,包括低样本大小对一些直径类和光线政权估计的准确性。然而,它也可能反映了现实。热带树种类似成人的身材和阴影公差可能会非常相似。
什么限制了树木生长?
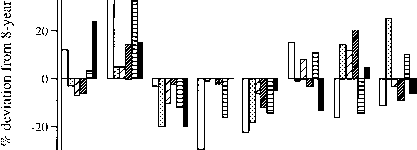
有趣的是考虑什么因素实际上限制个人的增长树在森林里。答案,正如已经指出,在大多数情况下是光的可用性。阴影效果在大多数树木在森林里增长速度远低于其潜力最大。克拉克&克拉克(1994)发现大型interyear增长变化的六种热带雨林,哥斯达黎加。在八年期间,直径增量为个人从50多厘米高1 cmdbh 3 - 10倍的最快增长较慢的(图3.9)。六种显示性能很强的一致性在观察期间,即站生产率是不同的相对统一。今年的最佳增长最低的一个降雨。似乎不可能穷湿年增长是由于浸饱水的土壤。更有可能是湿年多云雾,大幅降低光合有效辐射(PAR)收到干多年。
22个40
一年
图3.9年增长率平均直径变化的成年树(直径30厘米)的六个树冠和紧急在La热带雨林物种在原始森林,哥斯达黎加克拉克(Clark & 1994)。数据为每一个物种都是每年的年平均调整增量(平均超过所有个人在消除趋势或减去八年平均直径增加从每棵树的年度增量),作为百分偏差从物种的整体平均直径增加8年期间。横坐标标签显示的日历年开始每个测量周期。毫克,Minquartia glabra,洛杉矶,Lecythis ampla,嗯,Hymenolobium me-soamericanum, DP, Dipteryx panamensis, PE、Pithecellobium线虫,哈,Hyeronima alchorneoides。
在季节性干旱森林,增长可能减少由于缺水在旱季,除非树木能自来水在土壤深处。在La热带雨林、哥斯达黎加、直径增长的详细研究日常模式(Breitsprecher &伯特利1990)显示一个年度周期性增长大多数物种排水性良好的土壤。大多数物种显示轻度旱季减少增长在热带雨林。Barro科罗拉多岛上的1983年发生了一场严重的旱灾,有相当高的死亡率比正常情况下树,与大直径茎痛苦最大的增加死亡率。
食草性
材料的损失可以大量食草动物,即使在热带树木。完整的落叶树木经常被报道。此类事件很可能将树负面的性能影响。侯爵的研究(1984、1987)Piper arieianum述显示是这样的情况。研究小组在落叶的影响是下层木手掌(门多萨et al . 1987;Oyama &门多萨1990;Chazdon 1991;坎宁安1997)。大概是他们的大,相对较少,树叶简化观测。所有的热带棕榈研究证明是非常适应叶损失。 Indeed, Chamaedora tepejilote showed increased reproduction after being defoliated (Oyama & Mendoza
1990)。叶和无性系分株删除显示任何影响叶大小或茎直径Geonoma congesta落叶(Chazdon后三年
1991)。可能动员在缓冲存储的碳水化合物是重要的手掌从叶片过早脱落,但即便如此,坎宁安(1997)找不到任何显著影响存储碳水化合物的落叶Calyptrogyne ghiesbreghtiana。
继续阅读:其他的优势传播
这篇文章有用吗?