之间的相互作用的垂直混合和紫外线辐射的影响
前一节展示了混合过程确定方法UML成分(分子或生物)输入光敏区可能参与UVR-mediated过程。在强大的分层,这样的运输是非常有限的,所以紫外线辐射的影响只会涉及到这些选民已经出现在活跃区。如此极端的分层可以偶然重要系统昼夜斜温层下面的形式,但更通常的UML扩展下面水域中的光敏区,因此选民光敏区也将参与紫外线辐射光化学和photo-biology。UML的混合特征影响光学加工至少在两个基本方面。首先,UML的深度影响发生的平均紫外线照射;第二,垂直运输的速度,由于平流或湍流交换,决定了停留时间的光敏区(图2)。如果所有的生物和化学影响是累积暴露在所有时间尺度的线性函数,那么暴露的时间和顺序无关紧要的辐照度的垂直分布和知识(第三章)足以评估紫外线辐射的影响在水生生态系统。然而,大多数紫外线辐射效应是非线性的,至少在时间尺度的环境相关范围的一部分。的另一种方式说效果并不成比例累积暴露是互惠的失败。当互惠失败时,效果依赖于暴露的持续时间和辐照度,所以停留时间和平均辐照度产生影响。详细的例子,如何将这些依赖关系被认为是在下一节中出现。
4.3.1光化学
混合的影响光化学反应并不是众所周知的。作为光化学第一定律的一个推论(见章节6和8),主要的光化学反应速率应该是成正比的速度光吸收。然而,前面的声明只适用于完全混合,光学薄水体,或反应物的唯一吸收[39]。相反,在光学厚水列竞争在chromo-phores光子,混合率的差异可以影响光化学反应速率。在这种情况下,会释放CDOM流转很快从曲棍球池,这将转化为更高的光反应率[40]。
几个研究解决混合天然有机物的光降解的影响从大众运输的角度[40-42]。[40]me-socosm实验和模型(第四十一条、第四十二条)表明,混合利率可以施加巨大影响利率的光学加工和分销水柱。一般来说,缺乏,或缓慢搅拌范围光反应的顶端水柱水体中有强烈的光衰减的波长光反应和这些主要的光反应降低的利率随深度指数[41]。更彻底的混合,新鲜的光化学反应物不断添加到光敏层,导致较高的光解为整个水柱[43]。此外,高活性物种产生光化学地可能被垂直混合运送到更深的水域。例如,活性氧(ROS,第8章),产生表面附近可能经由混合和氧化有机分子几米在水面以下。
相反,缺乏(即混合。分层)也可能是重要的,因为它隔离一定CDOM池在UML分层期间。CDOM的UML(湖泊变温层)成为photobleached季节性暴露于紫外线造成的差异CDOM吸光度与表层之间更深的水域在湖泊(深水层)。例如,研究两个波科诺湖泊[12]发现光漂白的差异和紫外线辐射的光谱重量ing函数之间的CDOM漂白epilimnetic和hypolim-netic样本。这表明,分层的时机和程度可能影响photoproducts的年产量,包括碳气体,如二氧化碳和有限公司
前面的段落说明混合可以影响地表水的光化学反应的新建与新鲜,原生色团浓度和降低表面photoproducts [42]。但混合可能有额外的影响还未被探索。有机分子复杂的结构有几个光活化的网站(见第6章)。通过各种途径被吸收的能量衰减,这可能会或可能不会涉及分子重组和碎片[44]。在这种情况下,分子结构改变,由此产生的新结构可能有不同的吸收和photo-reactivity比其母分子特征。因此,除了沿着水柱影响大众运输,垂直循环可能会影响反应经历了由有机分子的序列。据我们所知,只有一个研究解决波动水平的太阳辐射的影响,在几分钟的时间尺度,天然有机物的光降解[45],(我)光漂白的研究显示差异,(ii)养分释放,和(3)后续使用CDOM的藻类和细菌,瓶子孵化之间在固定深度和瓶子旋转内水柱。虽然远未明确的,这样的结果表明,垂直混合应该明确地考虑在光化学研究的自然水域。
本节要求注意一些方式垂直混合复杂的光化学自然水域。另一方面,如果CDOM的利率吸收和光化学可以量化,然后稳态的光化学产品(如过氧化氢溶解)可以用来推断垂直混合率。这是可能的淡水系统(加拿大圣劳伦斯河和湖泊)积累较高的过氧化物由于其CDOM内容(36岁,46)。类似的尝试模型深时变化海洋中的过氧化氢(CDOM低得多的)只有部分成功繁殖观察到分布[47、48]。
4.3.2光生物学——浮游植物和bacterioplankton
浮游植物和bacterioplankton很小(0.2 -100鱼翅)单细胞生物体没有,或弱,能动性。一些bacterioplankton展览chemokinetic运动性,但是速度很小(s - 1(如0.001 - -0.01厘米。49])相比,典型的垂直运输率(1 - 10 cm s - 1的顺序)。浮游植物有不同属性的浮力和能动性与混合政权和交互影响光敏区停留时间(例子(16、18))。事实上,它一直认为藻类反映的各种生命形式的多样性混合政权水生环境[50]。在强烈搅拌(cf。图3),变异细胞的物理属性和能动性可能只有微妙影响垂直运输率但重要的垂直分布的影响,如积极或消极的浮力会导致
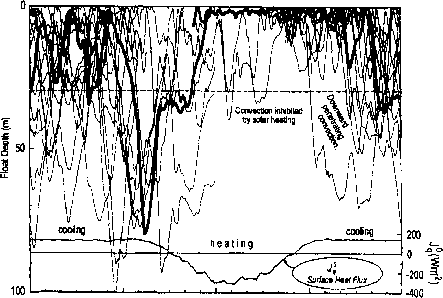
午夜中午午夜
每天的时间
图3。公布的漂浮的时间和深度分布示意图在不同时期在夜间的一昼夜的循环混合和拉布拉多海日分层(E。D 'Asaro和g Dairiki未发表的数据如2所示),这条沉重的线绳(复合三个独立的浮动部署)描绘了一个可能的深度的历史non-motile浮游生物在24小时期间。注意轨迹是终止一旦进入稳定昼夜变温层,因为他们真正开始提升由于轻微的积极的浮力漂浮。
午夜中午午夜
每天的时间
图3。公布的漂浮的时间和深度分布示意图在不同时期在夜间的一昼夜的循环混合和拉布拉多海日分层(E。D 'Asaro和g Dairiki未发表的数据如2所示),这条沉重的线绳(复合三个独立的浮动部署)描绘了一个可能的深度的历史non-motile浮游生物在24小时期间。注意轨迹是终止一旦进入稳定昼夜变温层,因为他们真正开始提升由于轻微的积极的浮力漂浮。
分布更大比例的浮游生物上或更低(分别)混合层的一部分。
浮游植物通过光合作用是一个重要的过程在水生环境和是众所周知的是与时间有关的。占垂直混合的影响作为一个行列式的历史是一个长期存在的问题的研究浮游植物生产力(见评论[2、14])。垂直混合可以影响生产力通过几种类型的时间响应[51];然而,整体光合作用时最常受影响的组合是由近地表辐照度敏感抑制[52]。
浮游植物对光线变化的时间尺度秒[51]许多天,平均曝光在一代人时间(ca。1 - 3天)有强烈影响抵抗的能力光抑制(参见第十章)。感应这些防御机制取决于之间的平衡PAR和浮游植物的新陈代谢的能量需求。如果平均光强度较低,浮游植物光合作用效率强调通过聚光色素积累和能力普遍薄弱防御标签暴露(PAR和紫外线辐射)。当这样的光线暗适应了浮游植物是运输在地表附近raybet雷竞技最新,它们可以严重影响标签暴露。另一方面,当光高可用性是相对于能量的要求,浮游植物强调保护和防御机制超过光合作用效率,积累保护性色素如叶黄素和UVR-screening化合物,和维护修理photodamage有效机制。
平均曝光的主要决定因素在混合层透明的水和UML的深度。因此,在相对稳定的水体光学特性,对紫外线辐射的敏感性会随着深度增加的混合,但混合深度防御机制应该是成反比关系。这个假说是由几个领域的研究。赫尔布等。[53]发现增加抑制光合作用的自然种群南极浮游植物增加深度的UML。而样本来自一个UML < 25米显示没有明显的抑制作用,抑制由于紫外线辐射约40%和75%的样本混合层深度35米,超过100米,分别。在他们的研究中,光合作用的抑制也减少了从南极到赤道,一部分是归因于UML的浅热带地区(参见图1),韦尔et al。[54]发现,紫外线辐射筛选化合物的浓度(一种适应标签暴露)呈负相关,UML光学深度(因此平均风险直接相关)南极半岛附近海域。尼尔等。[55]报道,对紫外线辐射的浮游植物Weddell-Scotia融合(南大洋)与表层密度有关,这是作为一个指标的整体光的历史。更高的深水表面密度反映了更大的贡献,从而更深层次的混合。混合深度似乎是一个重要因素解释在北方季节性变化的敏感性美国的五大湖[56],
大量的研究表明,快速的垂直混合可以提高生产如果表面接触很短的时间尺度的诱导时间相比,光抑制(年度)。另一方面,根据低垂直混合的昼夜变温层,光抑制非常明显(61、62)。影响其他指标的photoacclimation似乎也最大的时间尺度混合和光敏反应具有可比性(cf。63年,64]。这导致表达的一般观点,光抑制(票面价值或紫外线辐射),以长期孵化项目不适用的普遍条件下原位垂直混合,因此可以忽略(如。65]。
早期的研究生产力和垂直混合并没有特别考虑对紫外线辐射的反应,尽管一些早期,数量级计算表明它可能是重要的[66]。当然,在昼夜变温层的存在,表面扩展社区接受紫外线照射和严重抑制光合作用可能发生[36,67,68]。然而,另一个考虑是整体紫外线照射对水体的影响(垂直集成)生产。赫尔布等。[69]首先考虑垂直混合之间的关系和紫外线辐射影响综合生产使用的实验方法。屏幕旋转在石英管提供紫外线辐射和PAR梯度来模拟混合和获得的结果相比,平均汇率在静态条件下(无屏幕翻转)。孵化项目进行使用浮游植物在南大洋的大象岛附近的样本在屏幕覆盖管子的旋转速度估计的范围内运输在UML,紫外线辐射事件辐照度的100%和3%之间的不同。平均抑制浮游植物光合作用增加,当南极孵化的可变光政权(旋转屏幕)相比,一系列控制瓶,收到一个固定比例的入射辐射(图4)。在低平均辐照度的UML静态的浮游植物(即。碳固定,固定辐照度)低于样品在一个模拟的UML。在高辐照度,然而,样本“混合”的光合作用速率抑制(低)的平均值相比静态的治疗方法。
垂直混合可以提高抑制是一个令人惊讶的结果,实验表明,早些时候考虑垂直混合光抑制的影响降低了标准。然而,实验的浮游植物Weddell-Scotia融合(WSC,象岛附近)表明,光合反应动力学紫外线辐射非常不同于那些在早期的研究假设。光合作用发生迅速的减少后曝光,和恢复,在暴露于近地表辐照度,没有或减缓[55]。没有复苏意味着互惠的暴露应用short-time-scales但近地表足够强大,反应抑制非线性曝光超过30分钟
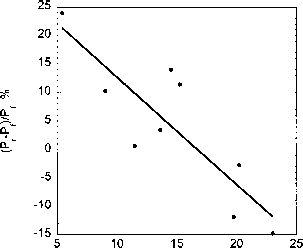
图4。综合孵化器主要生产12光水平(100%到3%的事件)当样品旋转(Pr)来模拟垂直运动在一个UML静态(固定)样本的百分之一(Pf)。变化百分比的函数给出总uv - b (290 - 320 nm, J m ~ 2)潜伏期。uv - b估计从测量辐照总量在320 nm (Em)测量比电磁建模应用到一个完整的紫外线辐射光谱的辐射传输模型计算晴空条件。(改编自赫尔et al . 69],
图4。综合孵化器主要生产12光水平(100%到3%的事件)当样品旋转(Pr)来模拟垂直运动在一个UML静态(固定)样本的百分之一(Pf)。变化百分比的函数给出总uv - b (290 - 320 nm, J m ~ 2)潜伏期。uv - b估计从测量辐照总量在320 nm (Em)测量比电磁建模应用到一个完整的紫外线辐射光谱的辐射传输模型计算晴空条件。(改编自赫尔et al . 69],
[55]。因此,高的光合作用并没有保持在短暂接触近地表辐照度作为标准影响被发现。这些不同的动力学的结果在数值模型研究不同紫外线照射的混合层的WSC [70]。类似于赫尔布等的实验结果。[69],垂直混合增强抑制集成水柱生产适度混合层深度(zmix < 40米)(图5)。在静态条件下,由紫外线辐射严重抑制,但仅限于表面附近的影响。根据光合作用随时间指数(或存活曲线,71][55]的关系。这些动力学的含义是,紫外线辐射影响最大的绝对在曝光时间的开始,即抑制发生在上半年的75%。因此,在静态层生产力的降低很小在第一次暴露在早上的两个小时。浮游植物的垂直混合导致通量从深度接触表层低到高的曝光度。在这种情况下,总是会有一个相对未曝光的组件表面的浮游植物将经历大,紫外线辐射诱导,光合速率降低。自恢复低(或不存在)之前暴露运输远离表层浮游植物,影响积累和综合抑制比强烈分层条件下高。 This situation has analogies with the mixing enhancement of CDOM bleaching as discussed in the previous section (4.3.1).
搅拌的速度不仅影响紫外线辐射的总体影响,还应对臭氧损耗(图5)。平流层臭氧损耗,这是
■•1 1。。 |
1 1 1 1 1 1 1 1 '我' ' 1 ' |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
v ^ |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- 10、30、60 |
\” |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
1分钟 |
你^ |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
1 ' 8 h: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
1 \ L, 4 h |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
——没有“”” |
1” |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
混合 |
\ 2 h: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
,,,我。。,我,。 |
T T续 T T续
0.90 0.95 1.00 0.90 0.95 1.00 图5。模型的结果对光合作用的抑制表层Weddell-Scotia融合的南大洋(修改尼尔et al . [70])。(a)的互动影响混合深度和混合时间尺度上每日水柱生产率相对于不羁率(曲线贴上imix =时间尺度混合)。朗缪尔循环可以混合上层水体迅速,所以混合搅拌时间< 1 h建模深处< 42 m。(b)的互动影响03损耗(150 DU vs 300 DU)和混合时间的函数混合深度:比例变化曲线一样的PT (a)。 最严重的极地地区,有一个幽灵似地具体影响太阳紫外线辐射光谱的uv - b辐射(280 - 315海里)尤其增强(第二章)。南极的计算效果“臭氧空洞”(即臭氧损耗50%)在水柱浮游植物生产静态情况下减少约1.5%,但多达8%减少混合场景[70]。同样,这似乎是光合作用的非线性响应的结果累积暴露。衰减的uv - b增加深度总是更快速衰减的紫外线a (WSC,和其他地方一样,图2也见第三章)。由于生物体暴露在紫外线辐射增强的臭氧损耗总是更大的表面附近。这个增量曝光导致更多的损失相对未曝光的浮游植物的光合作用运往大小写混合的表面相对于静态情况下,在增强的uv - b照射主要发生的近地表浮游植物光合作用的低税率甚至在正常uv - b的条件。 最近的实验进行了温和的巴塔哥尼亚海洋水域浮游植物的[72]还表示不同的反应在模拟混合紫外线辐射。(即垂直运动。辐照度的变化)和混合透光层深度(£ph值)是模拟使用移动系统结合中性密度屏幕,并与一个固定的系统。冬季和夏季的生产样品比较在相同的模拟的UML (unified modeling language, UML /£ph值< 0.5)。UVR-dependent减少碳固定利率下一个变量(“混合”)辐照度,物种适应低辐照度水平(即冬季样本,通常由microplankton)比夏天更敏感(由样品微型浮游生物)。此外,当夏天样本暴露在irradian-ces中遇到浅UML (unified modeling language, UML /£ph < 0.5),垂直混合增强紫外线辐射的抑制。然而,当深度模拟UML状态(UML /£ph > 0.8)对样品,在运动系统的浮游植物综合碳固定利率高于样品在固定系统。在这些研究中,动力学抑制依赖浮游植物种群的组成;有一个倾向于样品主要由微型浮游生物相对缓慢的抑制动力学。这也是观察研究不同安第斯湖泊[73],样品由较小的细胞抑制率比大细胞慢。 光合反应动力学明显是关键在预测垂直混合将如何影响紫外线辐射暴露的影响,到目前为止讨论的情况下。对比动力学已经观察到的浮游植物组合在其他温带环境。在这些组合中,复苏似乎更加活跃,因此互惠不是观察[74 - 77]。尽管抑制仍然是快速、稳态的速度获得经过短暂的过渡。这个稳态平衡的结果损伤和修复过程。当接触减少,光合活动恢复。当混合是足够长的时间尺度,建立稳态,紫外线辐射抑制光合作用仅是辐照度的函数,因此应该是独立的垂直混合率。然而,垂直混合的交互和紫外线辐射抑制光合作用已收到小实验检查组合活动修复。主动修复组合的一个例子是表层的浮游植物的温带瑞士湖(Vierwaldstattersee或琉森湖)[78]。波动的影响紫外线辐射在光合作用测定瓶流传的一个旋转升降系统通过一个表面混合层速度较快(完整的瓶子旋转每4分钟,曝光条件如图2所示)[79]。 The observed effect of UVR could be accounted for using a steady-state (irradiance dependent) relationship developed using laboratory incubations (Figure 6). Since the model of UVR effects is not time-dependent, this indicated that the effect of vertical mixing, if present, was too small to cause a deviation from the profile predicted under static conditions. While more studies are needed of how mixing interacts with UVR effects on phytoplankton production, the first indications are that results depend strongly on the kinetics of the UVR response.  图6。紫外线辐射对光合作用的影响(总C同化)的浮游植物通过不同的混合深度,每分光合作用表现为石英(紫外线辐射透明)相对于玻璃(部分紫外线辐射排除)瓶子。衡量利率的瓶子在表示深度范围流传的速度一旦每4分钟(0 - 2米),一旦每8分钟(0 - 3.9)和一次每20分钟(清廉和0 - 14米)4 h中午潜伏期。的建模利率平均稳态(基于辐照度)使用生物光合作用预测权重函数光合作用和辐照度(BWF /导出)曲线应用于原位辐照度估计从记录表面照度,瓶子和深度测量垂直消光系数。模型和测量在测量的可变性(ca。10%)同意除了清廉m孵化。实验在9月13日琉森湖,1999年9月15日,1999(没有星号)和(星号,看到暴露数据如图2所示)。(从科勒et al . 79年修改。) 图6。紫外线辐射对光合作用的影响(总C同化)的浮游植物通过不同的混合深度,每分光合作用表现为石英(紫外线辐射透明)相对于玻璃(部分紫外线辐射排除)瓶子。衡量利率的瓶子在表示深度范围流传的速度一旦每4分钟(0 - 2米),一旦每8分钟(0 - 3.9)和一次每20分钟(清廉和0 - 14米)4 h中午潜伏期。的建模利率平均稳态(基于辐照度)光合作用预测使用生物权重函数和光合作用辐照度(BWF /导出)曲线应用于原位辐照度估计从记录表面照度,瓶子和深度测量垂直消光系数。模型和测量在测量的可变性(ca。10%)同意除了清廉m孵化。实验在9月13日琉森湖,1999年9月15日,1999(没有星号)和(星号,看到暴露数据如图2所示)。(从科勒et al . 79年修改。) 生产由bacterioplankton也抑制UYR曝光(见[80]审查),但抑制之间的关系的深度和速度的垂直混合并没有被研究过。然而,细菌的DNA损伤的垂直分布,紫外线辐射效应的指标,研究了水列有不同的混合政体。最常见的UVR-me-diated破坏环丁烷嘧啶二聚体的形成(cpd)(见第9章)。有戏剧性的差异在垂直剖面cpd取决于平静的条件或强烈混合条件为准(图7)。Boelen et al .(81)观察到一个原位一昼夜的循环积累和减少表面cpd附近,而在平行表面孵化项目cpd增加中午下午但是没有修复。这表明垂直混合是必要交换严重破坏生物体(由于表面紫外线辐射暴露)的近地表层。    图7。测量和建模的环丁烷嘧啶二聚体(CPD)水列不同强度的垂直混合(a)墨西哥湾,1994年9月8日,混合层20米,风速低(2 ms-1) (b)墨西哥湾,1994年9月7日,混合层20 m,风速高(8 ms-1) (c) Gerlache海峡,南极洲,1996年10月6日,混合层25米,风速高女士(8”1)。建模概要文件仅供损伤(CPD)形成,攻击+光致复活作用,和损伤修复机制(光致复活作用和切除修复)。(Jeffrey等人与Huot et al . 82。) 图7。测量和建模的环丁烷嘧啶二聚体(CPD)水列不同强度的垂直混合(a)墨西哥湾,1994年9月8日,混合层20 m,风速较低(2 ms-1) (b)墨西哥湾,1994年9月7日,混合层20 m,风速高(8 ms-1) (c) Gerlache海峡,南极洲,1996年10月6日,混合层25米,风速高女士(8”1)。建模概要文件仅供损伤(CPD)形成,攻击+光致复活作用,和损伤修复机制(光致复活作用和切除修复)。(Jeffrey等人与Huot et al . 82。) 垂直剖面的细菌cpd反映的原位动态分层和混合,从而提供了一个机会来测试我们的了解垂直混合的过程,损伤和修复交互。这样的测试是由Huot等人建立一个数学模型的深度剖面的DNA损伤UML [82]。在这个模型中,DNA损伤的起始(cpd)是线性相关的暴露和DNA修复相关线性累积cpd利率从实验室研究开发。然而,photo-reactivation (PR)是任意减少到10%的响应预测从大肠杆菌中修复实验室测量为了DNA损伤的模型预测适合观察的概要文件(图7)。在这些假设下,混合率垂直分布的DNA二聚体,但几乎没有效果的影响平均二聚物浓度(82、83),作者认识到更多的数据需要在自然组合的细菌DNA修复之前现实建模可以做到的。率和线性都需要进行检查。观察破坏迅速修复时总cpd很低,但非常缓慢修复当cpd很高(Jeffrey等人[81],未发表),表明公关的相对有效性减少当损失超过一个阈值。如果是这样的话,平均损失在水柱会更高,当混合的速度放缓,CPD光敏区积累超过修复能力。 4.3.3水生生物——浮游动物andfish紫外线辐射的另一个重要的生物效应是多细胞死亡率增加水生生物,尤其是浮游动物和ichthyoplankton。至于浮游植物,浮游动物物种紫外线辐射的敏感性变化(第十二章),和敏感性的差异似乎主要是有关UVR-induced DNA损伤修复能力的差异(见第9章)和紫外线辐射的存在筛选化合物(第十章)。然而,一个重要的需要注意区分non-motile或缓慢的生物(例如,鱼卵)和活跃的游泳者,如甲壳类浮游动物和鱼类幼虫。前者可能被视为被动的成分,对紫外线辐射的平均在整个混合层,但后者的反应将更加复杂。大部分浮游动物可以长距离迁移垂直,现在变得明显的是,他们可以调整垂直位置紫外线辐射,甚至与紫外线(84 - 86,第14章)。这些浮游动物可能会寻求深躲避紫外线辐射的影响取决于他们浅或深的居住环境。 浅环境中的水垂直速度引起的湍流混合可能超过最大游泳速度的生物在任何深度的水柱。在平静的情况下,有机体可能无法达成紫外线辐射避难深水(取决于水的透明度和游泳速度),但在湍流条件下生物不可能避免被骑在整个水柱(cf。图3)。后者将强大的游泳者的唯一的有意义与平均生存在整个水体的垂直混合的速度。在上述情况下(浅和风),生物有效辐照度暴露对应的平均值在水柱和生物效应取决于有机体能够修复紫外线辐射的伤害。当修复缺失或低,互惠应用虽然之间存在非线性关系死亡率和曝光,要么作为一个简单的生存曲线(浮游植物,看到87年、88年)或更复杂的(但也非线性)对数曲线[89]。当修复,互惠不再听从和实际效果将取决于辐照度和暴露的时期。 这种类型的浅水环境中常见的阿根廷的巴塔哥尼亚草原和Pampean区域。湖泊浮游动物死亡率的一项研究发现在这些地区的存在与否互惠强烈影响数据从固定深度孵化项目是否可以用来预测生存在样品进行模拟混合(图8)。一个成功的物流关系的累积暴露在预测桡足动物的生存,Boeckella丁枫、缺乏photorecovery,桡足动物是否暴露在固定或旋转(不同的紫外线辐射)帧(89、90)。相比之下,另一个物种,Ceriodaphnia dubia,由光致复活作用能够修复DNA损伤,不服从相同的我们关系固定和可变(旋转)曝光。事实上,生存是更高的旋转样品相比,固定样本接收相同的累积暴露[90]。这些结果显示存在与否的互惠需要考虑在预测混合条件下浮游动物生存。 不同于以前的工作这些浮游动物对浮游植物生产力的研究没有直接解决这个问题的平均生存水柱是否受到垂直混合的速率的影响。这个问题更详细地检查了一对的缓慢移动的浮游动物对紫外线辐射的反应似乎遵守互惠:鸡蛋的大西洋鳕鱼(Gadus morhua)和胚胎的海洋桡足动物(Calanus finmarchicus)。对于这些浮游动物,生存是建模使用累积剂量的指数函数,类似于Weddell-Scotia光合作用的抑制融合(WSC)浮游植物(87、88)。类似的结果是获得的WSC浮游植物(参见4.3.2节):当平均水柱,生存是更高的低混合或静态情况下相比,快速混合[83]。类似的结果(增加平均存活率低混合)可能会对其它浮游动物缺乏修复机制(例如Boeckella丁枫)。然而,当修复是活跃的,垂直混合可能会增加平均生存在表层,所建议的Ceriodaphnia dubia案例已经讨论过(旋转样品最好生存比固定样品接收相同的剂量)。 在深环境中,强大的游泳者可以,原则上,逃离动荡的上层,避免有害的紫外线辐射暴露通过调整他们的垂直位置。因此,净UVR-induced死亡率可能是最小的。在这种情况下,成本,如果有的话,可能是间接的,即,由于紫外线辐射暴露和风险之间的平衡在表面水域捕食 100 80 40 60 0
继续阅读:理查德克泽普 这篇文章有用吗? 推荐项目 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
