太阳辐射调制器作为一个生态系统
罗伯特·g·吉姆
表的内容
摘要5
1.1介绍5
1.2规模很重要——辐射衰减与载荷的有机物6
1.3降水问题- 7频率和强度的影响的重要性
1.4直接影响紫外线辐射9
1.5移置与原地有机质-关键紫外可见介导过程调节异养利用9
1.5.1改变酶的可访问性的大分子10
1.5.2光解的腐殖质大分子10
1.5.3溶解有机氮和磷化合物的光解更正11
1.5.4完整的腐殖质物质的光解CO和CO2 11
1.5.5少直接但紫外线辐射的重要生物地球化学相互作用…12
1.6顽固的有机物质、代谢稳定性和光解13
引用15
文摘
太阳辐射是调制器的基本生态系统。几乎所有的有机物是光合作用,因此光的分布水生生态系统至关重要的监管主要能量输入。然而,同时特定组件的太阳能辐射,尤其是紫外线,函数作为一个加速器微生物降解提高生物利用度的复杂的有机基质微生物和组件的完整的光解和氧化的有机大分子二氧化碳和其他无机形式的营养。改变紫外线强度侵犯和内陆内水生和沿海海洋生态系统的自然或人为原因将修改的新陈代谢和生物地球化学过程与这些大分子相关。的级联效应可以大大修改自然生态系统的功能。
1.1介绍
本卷在随后的章节,详细评估提供一个汇总的当代理解紫外线(紫外线辐射)在水生生态系统的性质及其对水生生物的影响。我试图概述这些属性的耦合强调如何集成和个体紫外线辐射的影响,在生态系统层面,提供硕士级别的生态系统生物地球化学循环,调节能量通量、生产力,和系统进化。
关于这些特定组件的详细治疗太阳辐射的影响,强调的是很有用的几个相关的水生生态系统的普遍特征。生态系统是生物系统、生态系统生物地球化学系统、原材料和能源的循环生态系统是由一组高度可变的intercoupled物理、化学和生物参数。非常重要的是评估紫外线辐射的影响和改变生态环境的高度动态、变化的环境——动态空间,特别是时间尺度。然后的问题是紫外线的影响生态系统内的变量,分析混沌或某些化学计量分析允许定量预测的通用系统是否对紫外线辐射的变化的反应。
采取的方法是首先分析我们目前了解紫外线辐射的影响生态系统过程和这些过程如何intercoupled与其他相关这些过程的影响,如气候、大气过程与紫外线辐射有关。最后,可以合理地预测生态系统不同的特点将如何应对大气或水生条件的改变改变紫外线辐射。
1.2规模很重要——辐射衰减与载荷的有机物
几乎所有UV-C(< 280海里)被平流层吸收气体和水的水生生态系统。尽管相对较少的uv - b (280 - 320 nm)通过平流层(第二章),uv - b是高能和水域的重要photactivating代理。对紫外线a (320 - 400 nm)不如紫外线能量但不容易吸收和渗透水更深入。近紫外线蓝色部分的可见光谱(400 - 500 nm)最近已被证明是功能上类似于相邻a在许多重要的辐射光化学反应受紫外线辐射的影响,必须考虑任何评价的综合影响。
最近的原位测量了伟大的变化在uv - b的渗透和- a,但是普及率已经发现是远远大于先前被认为(3章6 [1])。当引用与纯水,传输的辐射大大降低了天然发色团的溶解有机化合物的浓度增加,特别是胡敏酸。紫外线衰减深度(表面irra-diance咱= 1%)范围从几厘米到> 10米的水域中[2 - 6],(> 90%)的among-habitat漫射衰减系数的变化(Kd)可以解释不同的溶解有机碳(DOC)的浓度。整个太阳能uv - b和a范围,Kd估计与一元权力模型是基于DOC浓度,特别是水域低到中度phytoplanktonic生产力。咱是强烈依赖于医生当浓度低于2毫克C 1 _1。在湖泊富营养化浮游植物的密度可以影响紫外线衰减[7]。
只有异构的某些部分溶解有机物(DOM)吸收太阳辐射。内河、酚醛等芳基腐殖质化合物(富啡和胡敏酸),很大程度上起源于陆生和水生植物,形成一个重要组成部分溶解有机酸和DOM可以构成一些总数的80%,其中30 - 40%是由芳香族碳化合物[8]。腐殖物质的最大组成部分载色体的溶解有机物(CDOM)。的可溶性部分腐殖质物质,异构富啡酸分子量从500年到1200年哒,包含许多酸性官能团,主要羧酸[9 - 11]。比富里酸和胡敏酸不亲水的大分子量(意味着ca。4000 - 5000 Da) [12]。腐殖质物质主导CDOM和最重要的组成部分,在吸收太阳紫外线和蓝色辐射(4,13)。
有机酸的浓度4 - 8毫克升1是常见的地表水,常常超过50毫克1 ~ 1在富含有机物的水域,如湿地、泛滥平原河流生态系统,间隙水hydrosoils [1]。CDOM和腐殖质物质的浓度通常沿着fresh-to-coastal-to-oceanic水域的梯度下降。
因为紫外线辐射对水生生态系统的影响是如此强烈受到CDOM的浓度的影响,因素的影响CDOM水生生态系统的加载速率会影响强烈紫外线的选择性分布及其对栖息地和生物的影响。在这方面两个方面尤为重要。首先,DOM的比例来源于高等植物组织(陆地和湿地/滨海资源)所主导的chromo-phoric腐殖质化合物与派生形式藻类,含有一些富啡和腐殖质组分(10,14、15)。小溪和河流的DOM是几乎完全由湿地高等植物和陆地的部分分解产品。同样的,小湖泊获得高比例的DOM从陆地和湿地资源主要由高等植物生产力和高比例的腐殖质物质残留部分退化的结构组织成分,特别是木质纤维素。
其次,接收水生生态系统的形态是必要的,因为直接湖盆体积水保留时间之间的关系,影响DOM的稀释,搅拌频率光区。大多数的数以百万计的湖泊很小(< 10平方公里)和相对较浅,通常在深度小于10米(1、16)。因此,交互DOM-entrained水与太阳辐射的频率通常是高的内部分层的湖泊在浅non-stratified湖泊和池塘。同样,DOM的频率在水中的小溪和河流与太阳辐射也很高,尤其是在更大的流订单(> 3日订单),从河岸树树冠阴影的影响很小。
1.3降水问题-频率和强度的影响的重要性
因为紫外线辐射的渗透和影响生态系统的新陈代谢和DOM功能是如此的强烈影响,DOM的装货率和时间接收水是很重要的。许多研究已经证明的主导地位移置输入的陆地有机物,碎屑DOM和颗粒有机质的形式(POM)和河流生态系统物质和能量循环流。大部分从土壤DOM被释放到地下水,毗邻湿地(如从厌氧过程。1,17 - 19)。
DOM输入从陆地有机物到河流和湖泊的结果直接浸出从生活植被和可溶性化合物在不同阶段进行径流从死去的植物材料的分解。非常高浓度的有机物来自湿地。输入的DOM通常直接与降水有关,与高加载率接收水域的初始冲洗阶段降水事件。DOM加载然后在后期随着稀释增加明显下降并最终排放体积下降。同样,DOM加载在最初阶段的融雪远远高于随后。尽管DOM加载高总在这些法拉盛事件,稀释也很高。最高的DOM浓度和产生的紫外线衰减发生时期低流成河。在分层湖泊,停留时间越长允许较高的光解的DOM的透光区。在浅,nonstrati-fied湖泊表面经常混合层的高紫外线日晒、水停留时间的集中效应的时间用于紫外线变更和微生物矿化(第四章)。
DOM加载的季节性时间也会影响UV光解和微生物利用的有效性。很明显,径流加载事件在寒冷,今年低光照时间将导致更有效的降解和利用生物有机化合物的生态系统。这些改变UV-mediated新陈代谢率会反过来影响营养再生和随后的许多生物生产力的水平。
DOM是交付给海洋沿海地区的河流,减少运输率发生在河口地区复杂的水动力弥散水流。低浓度的盐水覆盖的沿海水域和暴露在太阳能光解与强度和频率大于底层水域。结果是增加部分和完整的光解作用,主要由紫外线辐射,CDOM的矿化率较高的二氧化碳通过增强微生物的新陈代谢和二氧化碳的直接降解。结果,很大一部分剩余的DOM non-chromophoric (NCDOM)。也许这种相对顽固的NCDOM,构成总量的10 - 20% DOM,倾向于坚持与明显的化学稳定性和海洋环境长寿(几十年几百年)。
移置的加载率如何溶解有机物淡水生态系统和海洋地区和大陆将受到气候变化的影响尚不清楚。有迹象表明在长期数据集,在湖泊DOC浓度逐渐下降(如几十年。,l (p。779),20日,21日]。特别是在贫营养湖泊,DOC浓度通常较低,紫外线辐射穿透几米深的地方,可以负面影响生物体的遗传损伤,把生产转移到增加保护性色素的合成,或在高海拔和纬度高等植物源材料和DOM加载低。生物体在这样的湖泊可以暴露于高强度的紫外线辐射[22]。即使在湖泊DOM,含量更高的长期趋势往往对缓慢降低浓度的DOM [1]。
毫无疑问,这两个温度和大气中二氧化碳浓度增加。上升的温度也影响降水模式和导致大地区降雨和降雪积累减少[1]。干旱是大量的气象影响因素的累积结果降水、蒸散,和其他水的损失。干旱通常不会变得严重之前长时间的降雨不足和无节制的用水。
医生在一些湖泊在过去的四分之一世纪略微下降与实质性变暖(如重合。,l (p。780),23)。减少流域的降水和蒸散的增加导致减少流流动和降低文档加载到河流和湖泊。透明的湖水UV光解增加在这些条件下。类似的减少医生一直在观察流[19]。年度DOC收益率下降流发生尽管更高浓度在风暴流后长期干旱的时期(23、24)。
1.4直接影响紫外线辐射
藻类的光合作用显然是被暴露在自然水平的紫外线,特别是对紫外线a辐射。生理和遗传恢复时,结果发现准稳定的生理状态之间的一般损伤和恢复过程[25 26章9、11和13]。许多物种光和DNA损伤修复日常时期的黑暗。许多物种产生防紫外线化合物——mycosporine-like氨基酸是一个重要的和无处不在的类的化合物(27日28日,第十章)。许多物种的生化防御有毒紫外线辐射的终端产品,如自由基清除由类胡萝卜素色素和超氧化物歧化酶(第15章)。一些物种有限能力避免强烈的紫外线表面迁移到更深的领域。
紫外线辐射会影响浮游动物和鱼类直接在浅水栖息地破坏DNA和一代的有害的光化学(自由基活性氧)(29、30、第8章)。尽管许多动物可以避免UV-intense栖息地,以及开发光保护颜料(类胡萝卜素,表皮黑色素),这两种策略可以改变他们对捕食其他生物,特别是鱼。
1.5移置与原地有机质-关键紫外可见介导过程调节异养的利用率
约90%或更多的水生生态系统的总代谢是微生物,通过异养代谢细菌,真菌,和许多原生生物,所有大小小于100¡害怕[1,31日]。因此,水生生态系统的物质和能量通量是完全由代谢颗粒碎屑(非生物),特别是从原地和移置来源DOM。远洋打开水,但整个湖泊或河流生态系统的一部分。与DOM加载和通量,移置和滨海来源是至关重要的,因为它们的化学区别产生的藻类光合作用。
衰老的模式、死亡和降解率的生物群也相当重视利率和通路的退化和能量利用率。例如,持续减缓衰老和释放的DOM高等水生植物非常不同于DOM的相对瞬时生化死亡和释放细菌和藻类。Non-predatory死亡和新陈代谢的非生物碎屑POM和DOM通过原核和原生生物的异养生物主宰所有水生生态系统。
提供合成紫外线对水生生态系统的影响,一个关键组件是同时DOM的重要性在调节的分布和衰减UYR以及紫外线的影响具有直接和间接的新陈代谢、生长、生殖、和生产生物群的功效。因为这些影响紫外线交互和耦合,很难分开没有冗余。几点可以为特征,然而,在总结一些更详细的讨论在后续章节。
物理过程,如部分或完整的光化学修饰有机大分子,可能导致重大改变生物可用性的部分复杂的、异构的溶解的有机化合物。这些光化学过程会导致:
1.5.1改变酶的可访问性的大分子
发生在伟大的丰富多酚有机酸(通常4 - 8毫克1 _1)在许多新鲜的水,可以用或诱导沉淀蛋白质的复杂绑定到一个或多个网站蛋白质亲水表面产生一个单层小于蛋白质本身(32、33)。这种络合,以及交联多肽链的多酚腐殖质物质会导致酶的抑制或减少活动(如。,到三十五]。更多的芳香和浓缩胡敏酸分子更严格,能更大程度上扭曲结合酶比是这样的简单的化合物,如富里酸(如。,36)。酶的抑制作用发生在古典非竞争性的方式,抑制剂,多酚,同时底物结合到酶。此外,溶解腐殖质物质可以通过peptidization复杂和改变生物对酶法水解。例如,膜性质,如脂质疏水性,可以改变腐殖质物质,进而影响酶水解率和营养传输机制(如。、37、38),生态系统一个重要的方面是这些蛋白质或酶复合物可以存储在一个长期灭活状态,运输在生态系统内,后来重新激活部分由紫外线辐射光解的乳沟[1,34岁,35]。
1.5.2腐殖质大分子的光解
部分光解腐殖质大分子,特别是挥发性脂肪酸和相关的简单化合物的生成,作为优秀的基质细菌降解(如。,39-42]。总额的重要的是要认识到,光解的辐照度,大约四分之一的部分光解的有机基质uv - b的结果,大约有一半来自- a,并从低四分之一波长(400 - 500 nm)光合有效辐射(PAR, 400 - 700 nm)。透光率和光解的活动从uv - b和a主要局限于表层海水。相比之下,不相上下,尽管比UY大力弱得多,更大的深度渗透到水。虽然有机化合物的光解作用明显小于由紫外线引起地表水,光解的一代的简单的底物是明显的由标准以及紫外线[1,43岁的44],这些研究结果表明一个可观的一部分光解的一代的一些简单的基板是由标准。
1.5.3光解更正溶解有机氮和磷的化合物
光解的退化的溶解有机氮和磷化合物释放无机营养物质如亚硝酸盐、氨、CO和CO2和磷酸盐以及(如。,回顾41,45-47]。显然刺激增加养分有效性的影响这些过程发生(如。,47]。
1.5.4完整的腐殖质物质的光解CO和CO2
光化学氧化由太阳辐射的自然溶解有机化合物有限公司和溶解无机碳(二氧化碳和HC03 ~)已经知道多年(如。,48]。根据离解和饱和条件下,一些多余的二氧化碳会逃避到大气中。先前的研究光解的降解的溶解有机物表明,太阳辐射的主要光解的组件是uv - b和a,,一样的后果很小。然而,许多这些研究没有在无菌条件下进行,结果发现感到困惑几乎瞬时产生的有机化合物微生物利用快速降解和生成的二氧化碳。此外,许多的DOM来源这些研究已经长时间暴露在电离辐射(如周)和消逝的光。现代研究表明尽管uv - b和a意义重大,可以导致超过一半的光化学矿化,标准也是一个主要光解的代理(43,49岁,50)。例如,来自近200个单独的光解的实验DOM不同水域和植物在不同条件下,频谱的uv - b部分总是最有效的完整的光降解为二氧化碳,但对紫外线a也是非常有效的小差异从紫外线的光解的能力[1]。PAR也高效光解的DOM co2退化,经常约四分之一到一半的集体光解作用可以在很大程度上归因于蓝色部分的频谱相媲美。CDOM的生物利用度可能增加[40],仍然没有改变,或减少从光解[47,51岁,52]。生物利用度明显相关阶段的光解和变更的主要组件的异构自然聚合的天然有机化合物。
两部分光解产生的挥发性脂肪酸,和完整的光解产生的大量二氧化碳水平是重要的发现,因为票面价值的物种灭绝率低得多的水相比,紫外线辐射。光解的过程,所以重要的养分循环,因此不局限于几厘米的地层的水生生态系统,而是影响大部分seasonally-variable透光层的体积。
1.5.5减少紫外线辐射的直接而重要的生物地球化学相互作用
生物地球化学紫外线辐射的相互作用在DOM水生系统也很重要,但缺乏研究生态系统水平。继续深入研究水生生态系统的自然溶解有机物质是导致改进的理解这些不同化合物的许多方面,尤其是腐殖质化合物,可以与其他重要代谢组件交互。这些过程会改变紫外线部分或完整的DOM光解。例子是多方面的:
(一)相互作用无机化合物,尤其是在络合反应如螯合了53。根据浓度比率的络合DOM无机元素、有机络合的模式生物可用性,在某些情况下,元素可以增加或减少毒性。所有的这些过程将由UV光解改变ofCDOM。
与其他有机化合物(b)进行交互,如peptidization和改变生物对酶法水解。例如,膜性质,如脂质疏水性,可以改变腐殖质物质,进而影响酶水解率和营养传输机制(如。、37、38)。最有趣的互动,腐殖质物质能与蛋白质复杂,尤其是自由可溶性和膜结合酶与非竞争性抑制[54、55]。酶可以存储长时间(数天、数周)在这个复杂的,不活跃的状态,在与水生态系统重新分配包裹运动,并重新激活部分由紫外线辐射光解的劈理(31岁,34岁,35岁,43)。
(c)改变氧化还原等化学性质和博士的优势胡敏酸会导致有机酸性能的影响,有时超过,及派生的酸度形成自然或人为来源[综述1]。接触自然溶解有机物的紫外线可以形成减少活性氧物种,特别是过氧化氢(H202) [56、57;第8章)。H202半衰期的几个小时在自然水域,可以从根本上改变金属的氧化还原循环[58]。
(d)微生物降低腐殖物质可以进入少减少区域的沉积物,作为电子给体的微生物还原几个环境重要的电子给体[59]。一旦微生物减少,腐殖质物质可以电子转移到各种铁(m)或锰氧化(iv)形式abiotically和回收腐殖质氧化形成化合物,它可以接受更多的电子从腐殖质compound-reducing微生物。这些高活性的紫外线辐射的相互作用过程浅水区,特别是滨海湿地地区,尚不清楚,(e)改变选择性修改等物理性质光穿透。著名的选择性的光衰减CDOM (cf。1)在很多方面可以进一步修改生物地球化学循环。这样修改的光合作用的光政权可以改变利率,荷尔蒙活动,迁徙分布和生殖行为。吸收的UYR腐殖质物质可以保护生物体免受基因损害以及修改大分子,提高生物利用度的有机基质。
1.6顽固的有机物质、代谢稳定性和光解
医生的普遍观察到的不完整的光解加速利用这些大分子至关重要,但显然不是强制性的。部分复杂DOM池,包括分数腐殖酸和富里酸化合物,是退化,但总降解率明显放缓。化学有机顽抗的DOM有助于提供一个热力学稳定的新陈代谢在湖泊,水库,wetland-littoral地面水达到界面,河流生态系统[1,31岁,54岁,60 - 62]。化学固执是一种“刹车”生态系统的新陈代谢,刹车是集成的关键维护稳定的异养利用合成或进口有机物质和能量。U虚拟现实可以改变化学顽抗的有效性“刹车”。
大多数的有机碎屑池,在微粒和溶解阶段,内陆的水生生态系统由残余有机化合物的植物结构组织。越不稳定的有机成分复杂的溶解和颗粒有机物通常水解和代谢速度比少访问en-zymatically顽固的有机化合物。结果是一个一般的顽固化合物浓度的增加,通常超过总数的80%,慢的代谢率和周转率。这些顽固的化合物,然而,代谢速度减缓和监管在很大程度上由其分子复杂性和焊接结构。
在每一个详细的年度湖和河流生态系统有机碳预算,有机质生成的浮游植物不会支持所有的异养代谢的生态系统。至少几倍的总代谢支持有机地面水达到界面的补贴社区和移置生产。从代谢稳定性的角度来说,它是特别重要的,大多数有机碳的溶解和相对顽固的,广泛分布在内陆水域。化学顽抗的主导改善着暴力的代谢和生长的DOM振荡的特点深海生物群生态系统的组件当资源丰富。此外,大部分的POM主导地面水达到界面区域中形成的湖泊,河流,和estuar-ine生态系统,是减少流离失所,缺氧的环境滨海和深海底沉积物。医生,主要是高等植物的起源,提供了定量的货币稳定和碎屑在水生生态系统途径更重要。相同的基础,稳定在陆地生态系统,可能在沿海以及如果不是大部分的海洋生态系统。
碎屑包括非生物微粒、胶体和溶解有机物,新陈代谢率大小只影响水解攻击[31]。内陆水生生态系统收集有机物质,特别是在溶解的形式,从陆地,湿地和沿海资源的数量,补充如果不超过这些产生了原地。有机物的利用是降低化学顽抗的结合以及位移缺氧环境。因此,内陆水生生态系统是异养和功能碎屑碗,不是藻碗。
地面的高有机物生产尤其是地面水达到界面区域(湿地、沿海地区)通常导致加载过多的有机碳,通常主要是溶解有机化合物的形式,内陆水域。很大一部分,DOM是代谢,这些纤维和沉淀,或photolyzed移动时通过湖泊和河流,但部分到达海岸海洋区域。这移置加载海洋水域的程度尚不清楚,但估计高达总数的20%海洋DOC (63、64)。由于长时间的接触多的移置DOM光解的降解过程途中开放海域,其代谢监管职能明显小于在内陆水生态系统。更不稳定的DOM海洋深海藻类光合作用占主导地位的产品,结果进行快速的利用和开发,直到持续增长限制条件为准。假设这些条件明显不稳定,这些含有高浓度的发色团的和non-chromophoric DOM所主要来自高等植物组织。因此,改变利率的影响紫外线辐射的影响海洋深海生态系统的影响更直接的手段,如直接破坏生物的遗传成分,而不是改变有机大分子的化学的主要角色。
在长期进化的尺度,人类现在有能力迅速干预这种相互依存关系和稳定性的改变有机物的代谢率。例如,减少平流层的臭氧和相关的uv - b增加可能导致加速光解的降解大分子的DOM co2非生物和生物通路。此外,对细菌代谢基质的光解的增强UV光解可能导致加速养分生物地球化学循环和刺激生产力的生态系统。除了减少湖泊和溪流的代谢稳定性,增强微生物呼吸肯定会导致一代向大气中二氧化碳和逃税。
引用
1。R.G.吉姆(2001)。湖沼学:湖和河流生态系统,学术出版社,圣地亚哥。
2。J.T.O.柯克(1994)。光学在自然水域的uv - b辐射。拱门。Hydrobiol。Beih。Ergebn。Limnol。43岁的论文。
3所示。新墨西哥州史卡利,D.R.S.瘦(1994)。紫外线辐射的衰减在温带湖泊。拱门。Hydrobiol。Beih。Ergebn。Limnol。43岁的35-44。
4所示。D.P.莫里斯h . Zagarese石球威廉姆森,例如Balseiro,开国元勋之一B.R.安贝德卡对哈格里夫斯,b . Modenutti r·穆勒c Queimalinos (1995)。太阳紫外线辐射的衰减在湖泊和溶解有机碳的作用。Limnol。Oceanogr40,1381 - 1391。
5。石球威廉姆森,R.S. Stemberger D.P.莫里斯T.M.霜,密度Paulsen (1996)。紫外线辐射在北美湖泊:衰减估计医生测量和对浮游生物群落的影响。Limnol。Oceanogr41岁,1024 - 1034。
6。r . Sommaruga r . Psenner (1997)。紫外线辐射在奥地利阿尔卑斯山的一个高山湖泊:空中和水下测量。Photochem。Photobiol。,65,957 - 963。
7所示。渡边y, y Hodoki (1998)。太阳紫外辐射的衰减eu-trophic淡水湖泊和池塘。日本。59岁的j . Limnol 27-37。
8。R.L.马尔科姆(1990)。每个土壤腐殖物质的独特性,河流和海洋环境。肛交。化学。学报,232,19-30。
9。e . Saski a . Vahatalo k . Salonen m . Salkinoja-Salonen (1996)。Mesocosm模拟沉积物形成引起的生物漂白硫酸盐纸浆厂废水处理在淡水中。:m .伺服k . Munkittrick j·凯里,g .从克拉克(Eds)。、环境命运和纸浆和造纸厂废水的影响,(261 - 270页。)圣露西出版社,德尔雷海滩,FL。
10。D.M.麦克奈特,广义相对论艾肯(1998)。源和水生腐殖质的时代。:交货单黑森州,l·j·特兰威克(Eds)。水生腐殖质物质:生态和生物地球化学。(页9-39)。斯普林格出版社,纽约。
11。j . Peuravuori k Pihlaja (1999)。描述水生腐殖质物质。:j . Keskitalo p Eloranta (Eds)、(零部件)页。湖沼学的腐殖质的水域。Backhuys出版商、莱顿、荷兰。
12。S.E.卡巴尼斯、问:周Y.-P P.A.莫里斯。下巴,广义相对论艾肯(2000)。对数正态分布分布模型水生富里酸的分子量。环绕。科学。抛光工艺,34岁,1103 - 1109。
13。通用汽车法拉利,医学博士道(1998)。CDOM吸收特征与荧光和盐度南部沿海地区的波罗的海。河口、沿海大陆架科学。,91 - 105。
14。D.M.麦克奈特,广义相对论艾肯,R.L.史密斯(1991)。水生富啡酸在基于微生物的生态系统:从两个结果南极沙漠湖泊。Limnol。Oceanogr。,36岁,998 - 1006。
15。D.M.麦克奈特,既有安德鲁斯R.L.史密斯,r . Dufford (1994)。水生富啡酸在南极algal-rich池塘。Limnol。Oceanogr。,39岁,1972 - 1979。
16。R.G.吉姆(1990)。地面水达到接口:代谢和湖沼学的监管机构。Verhand。国际的。联盟。Limnol。6日到24日,24日。
17所示。灯光。达姆,再见Trotter jr Sedell (1987)。厌氧区和流程流的作用生态系统生产力。直Averett): D.M.麦克奈特(Eds)。、化学
水质量和Hydrologie周期(页157 - 178)刘易斯出版商,切尔西,MI。
18岁。西奥多B.W.堪称摩尔(1990)。控制溶解有机碳浓度流,魁北克南部。可以。j .鱼。Aquat。Sei。,1537 - 1544。
19所示。P.J.狄龙,洛杉矶Molot (1997)。影响景观形式出口的溶解有机碳、铁、从森林溪流集雨和磷。水Resour。Res, 2591 - 2600。
20.D.W.辛德勒卢铁荣霜K.H.米尔斯,P.S.S. Chang I.J.戴维斯D.L.马利测向,芬德雷J.A.希勒,M.J. Turner pi Brezonik, a·斯文森(1991)。对比实验和atmospherically-acidified湖泊在压力和恢复。Proc。皇家Soc。爱丁堡,97 b, 193 - 226。
21。D.W.辛德勒,P.J.柯蒂斯(1997)。医生的角色在保护的相似性(原文如此)受到紫外线照射的气候变暖和酸化。生物地球化学,36岁,1 - 8。
22。D.M.麦克奈特,r·尼斯R.L. Wershaw, j.s男爵,s·希夫(1997)。微粒的化学特性、胶体和溶解有机物在尼斯淡水河谷的分水岭。生物地球化学,36岁,99 - 124。
23。D.W.辛德勒,贝利S.E. P.J.柯蒂斯,开国元勋之一B.R.安贝德卡对帕克斯塔特,议员c.a凯利(1992)。自然和人为影响因素溶解有机物质的富足和骑自行车前寒武纪盾湖泊。229年Hydrobiologia, 21
24。S.L.希夫M.J.辛顿说,贝拉英语(1997)。风暴的浓度和出口的重要性从两个前寒武纪盾集雨溶解有机碳。生物地球化学,36岁,67 - 88。
25。J.J.卡伦,P.J.尼尔(1994)。紫外线辐射、臭氧损耗、和海洋光合作用。Photosyn。Res, 39岁,303 - 320。
26岁。d . Karentz马丁博思韦尔,R.B.棺材,a·汉森GJ。Herndl, Kilham轮议员较小,m . Lindell右眼Moeller, P.J.尼尔,前作空。桑德斯,C.S.维勒,R.G.吉姆(1994)。uv - b辐射对远洋淡水生态系统的影响。拱门。Hydrobiol。Beih。Ergebn。Limnol。,43岁,31 - 69。
27。W.F.文森特·s·罗伊(1993)。太阳能紫外线b辐射和水生主要生产:破坏,保护和恢复。环绕。牧师,1,1 - 12。
28。熊,j . Komenda l . j . Kopecky Nedbal (1997)。微小的藻类的紫外线防护策略。杂志。工厂。,100,378 - 388。
29。交货单黑森州(1994)。水蚤对紫外线的反应。拱门。Hydrobiol。Beih。Ergebn。Limnol。,43185 - 195。
30.o . Siebeck T.L.维尔,石球威廉姆森,r .检查者,d .黑森州h . Zagarese大肠,大肠Balseiro, b . Modenutti j . Seva a .家住(1994)。uv - b辐射的影响在远洋淡水生态系统浮游动物和鱼类。拱门。Hydrobiol。Beih。Ergebn。Limnol。,43岁,101 - 114。
31日。R.G.吉姆(1995)。死亡、碎屑和水生生态系统的能量流。Freshwat。医学杂志。,33岁,83 - 89。
32。e·海斯蓝(1988)。植物多酚(syn.植物单宁)和化学防御——重新评价。j .化学。生态。,1789 - 1805。
33。e·海斯蓝(1988)。实际的多酚:从结构到分子识别和生理作用,剑桥大学出版社,剑桥。
34。R.G.吉姆(1991)。细胞外酶在水生生态系统相互作用:存储、再分配和种间的沟通。:R.J. Chrost(主编),微生物酶水生环境,6-28 (pp)。斯普林格出版社,纽约
35。蔡明俊。Boavida, R.G.吉姆(1998)。抑制磷酸酶活性和溶解腐殖质物质的水解复活自然紫外线。Freshwat。医学杂志,285 - 293。
36。J.N. Ladd,巴特勒J.H.A. (1975)。Humus-enzyme系统和合成,有机polymer-enzyme类似物。保罗:电子艺界,公元迈凯轮,(Eds)、土壤生物化学(4卷,第194 - 142页)。m·德克尔公司,纽约。
37岁。功率因数M.J. Lemke说丘吉尔,R.G.吉姆(1995)。基质和细胞表面疏水性的影响在细菌磷肥利用率。达成。环绕。Microbiol, 61, 913 - 919。
38。功率因数M.J. Lemke说丘吉尔,R.G.吉姆(1998)。胡敏酸与细胞外的交互层湿地细菌。Verh。国际的。联盟。Limnol26,1621 - 1624。
39岁。A.J.斯图尔特,R.G.吉姆(1981)。溶解腐殖质材料:光降解、泥沙的影响和反应性磷和钙碳酸盐沉淀。拱门。Hydrobiol。,92,265 - 286。
40。R.G.吉姆,P.G.孵卵,T.S.比安奇(1995)。紫外辐照度自然光解的顽固的溶解有机物为快速简单的底物细菌的新陈代谢。Limnol。Oceanogr。,1369 - 1380。
41岁。硕士Moran R.G.泽普(1997)。光反应在生理不稳定化合物的溶解有机物的形成。Limnol。Oceanogr。,42岁,1307 - 1316。
42。硕士Moran j.s秘密。Photochemically-mediated溶解有机物与bacterioplankton之间的联系。:s . r . Sinsabaugh芬德雷(Eds)、集成方法Microbial-Dissolved有机质营养联系,学术出版社,圣地亚哥,在新闻。
43。R.G.吉姆(2002)。起源、命运和自然湿地有机化合物的影响。:M.M.荷兰,马丁·沃伦,正当Stanturf (Eds)、湿地和可持续性水资源将军科技,众议员SRS-50森林服务,(页183 - 189),美国农业部Ashevill,数控。
44岁。R.G.吉姆,北卡罗来纳州Tuchman,影响co2浓缩植物降解产物的生产及其自然的光降解和生物利用率(2002)。Limnol。Oceanogr。47岁的审查。
45岁。文学士学位曼尼,贝拉米勒R.G.吉姆(1971)。紫外线燃烧湖水域中溶解有机氮的化合物。Limnol。Oceanogr。,71 - 85。
46岁。R.J. Kieber, a . Li P.J. Seaton (1999)。生产的光降解亚硝酸盐溶解有机物在自然水域。环绕。Sei。抛光工艺,33993 - 998。
47岁。A.V. Vahatalo, k Salonen,明斯特·m·贾维R.G.吉姆(2002)。光化学转换移置有机物提供了可利用的营养的腐殖质湖。拱门。Hydrobiol。在出版社。
48。朱柔米勒R.G.泽普(1995)。光化学的生产从陆地有机物溶解无机碳:意义海洋有机碳循环。地球物理学。卷。,22岁,417 - 420。
49。A.V. Vahatalo, m . Salkinoja-Salonen p . Taalas k Salonen (2000)。光谱光化学量子产量的溶解有机碳的矿化腐殖质湖。Limnol。Oceanogr。,45岁,664 - 676。
50。R.G.吉姆(2000)。自然uv - b光降解的溶解有机物的分解植物来源容易降解脂肪酸。Verband。Int,联盟。Limnol, 27岁,2036 - 2043。
51。点Anesio, Denward, l·j·特兰威克,w . Graneli (1999)。减少细菌生长在维管植物碎屑由于光化学修改。Aquat。活细胞。生态,17159 - 165。
52岁。Obernosterer, b . Reitner G.J. Herndl (1999)。对比太阳辐射对海洋bacterioplankton溶解有机物和生物利用度。Limnol。Oceanogr, 44岁,1645 - 1654。
53岁。E.M.濮德培(1998)。化学成分、结构和金属绑定属性。:交货单黑森州,l·j·特兰威克(Eds)、水生腐殖质物质:生态和Bio-geochemistry(41 - 61页。)。斯普林格出版社,柏林。
54。R.G.吉姆(1992)。Gradient-dominated生态系统:溶解有机质的来源和监管职能淡水生态系统。Hydrobiologia, 229, 181 - 198。
55。R.G.吉姆(1993)。从湿地腐殖质化合物:络合、失活和活化表面束缚和胞外酶。Verhand。国际的。联盟。Limnol。,25122 - 128。
56。j•库珀R.G. Zika病毒,R.G. Petasne, J.M.C.平面(1988)。光化学的形成在自然水域H202暴露在阳光下。环绕。科学。抛光工艺,22岁,1156 - 1160。
57。新墨西哥州史卡利,D.R.S.精益D.J.麦昆j•库珀(1995)。光化学过氧化氢在湖泊的形成:溶解有机碳和紫外线辐射的影响。可以。j .鱼。Aquat。科学。,52岁,2675 - 2681。
58岁。J.W. Moffet, R.G. Zika病毒(1987年)。过氧化氢反应动力学与铜和铁在海水中。环绕。科学。抛光工艺。21岁,804 - 810。
59。湄Lovley J.L. Fraga,一口油井为el瑞Blunt-Harris J.D.科茨,(1999)。腐殖质作为厌氧呼吸的电子供体。环绕。Microbiol。,89 - 98。
60。R.G.吉姆(1983)。湖沼学(第二版)。桑德斯学院出版、费城、
61年。R.G.吉姆(1984)。碎屑和颗粒溶解有机碳在水生生态系统功能。公牛。3月科学。,35岁,503 - 509。
62年。R.G.吉姆(2000)。淡水生态学:需求变化,未来的需求。湖沼学1,3-11。
63年。m . Meybeck (1993)。河边的大气碳运输:来源,全球类型学和预算。水空气土壤调查。,70443 - 463。
64年。m . Meybeck (1993)。天然来源的C, n . P和S: r . Wolast F.T. Mackenzie, l·周(Eds)交互的C, n . P和S生物地球化学循环和全球变化(页163 - 193)。斯普林格出版社,柏林。

物理
•紫外线辐射气候学
•水体光学和渗透的紫外线辐射
•调制垂直混合的紫外线照射和效果,平流
第二章
紫外线辐射气候学
马里奥Blumthaler和安·r·韦伯
表的内容
抽象的23
2.1介绍23
2.2理论24
2.2.1能源来自太阳的24
2.2.2行星运动和几何25
2.2.3大气27
2.2.4吸收和散射29日
2.2.5确定紫外光谱在地上33
2.2.5.1臭氧吸收33
2.2.5.2平流层臭氧化学34
2.2.5.3 35平流层臭氧的变化
2.2.5.4对流层臭氧化学35
2.2.5.5其他衰减器36
表面37 2.2.5.6最终结果
2.3测量39
2.3.1地面测量39
2.3.1.1仪表39
2.3.1.2结果42
2.3.2 Space-born测量50
2.4太阳紫外线辐射52的趋势
2.4.1长期52臭氧变化
2.4.2长期紫外线辐射变化52
2.4.3未来的紫外线辐射水平54
2.4.3.1预测紫外线辐射54
2.4.3.2未来紫外线场景54
引用55
文摘
紫外线辐射在地球表面是由太阳的辐射,穿过大气层时和随后的修改。地球到太阳距离的和地球上的观察者的位置确定入射辐射的力量而在地面的紫外光谱随时间和地点,由于波长的依赖在大气衰减过程。最重要的决定因素是太阳仰角、臭氧和气溶胶内容,高度和反照率和阴沉。测量的高光谱分辨率允许详细调查的这些参数的影响。简单的宽带测量从大量的位置提供信息。模型的计算可以估算辐照度水平如果输入参数(大气)是众所周知的。因此,估计基于space-born测量提供全球分销和时态变化紫外线辐射,但不完整的知识一些大气参数仍然限制了绝对精度。
总臭氧量减少,尤其在中期和中高纬度大陆地区,uv - b增加。所有潜在的气候变化之间的相互作用影响臭氧使得预测未来紫外线的困难,raybet雷竞技最新但是不要期望恢复在全球范围内的最佳估计早于在10到20年。
2.1介绍
紫外线辐射进入水生生物在自然栖息地来自太阳。额外的,地面辐射修改穿过地球大气层,有许多因素影响辐射到达地球表面。其中包括大气的状态(清晰、干净、多云、污染),地球上的位置(纬度和海拔)和季节(相对太阳的位置地球上的位置)。进一步衰减然后发生辐射穿过水环境达到水生生物。这些在第三章后并发症处理;这里我们只处理地面紫外线辐射事件,或水的表面。
紫外线辐射覆盖的部分电磁波谱在波长400纳米以下,x射线和可见辐射。紫外线是分裂,有些随意,为窄波段名称(从不同的分支科学),如真空、远、近紫外线。在考虑紫外线在地球表面,我们关心的是最长的波长紫外光谱的一部分,那些在280 - 400海里,称为对紫外线a和紫外线辐射(更有害UV-C (200 - 280 nm)和更短的波长是完全由大气衰减)。委员会国际歌d 'Eclairage (CIE) uv - b定义为280 - 315纳米,和对紫外线a 315 - 400 nm。然而,uv - b经常可以发现描述为280 - 320纳米,务实的原因,320海里的地方太阳光谱“筋疲力尽”,生物的地方行动光谱方法一个地区零或非常小的反应。在现实中这些波段的区别是任意绑定白羊座在连续光谱的太阳辐射和生物反应。
本章的第一部分探讨了辐射和辐射传输的基本物理。在下面几节中,讨论了紫外线辐射的测量,在地面和space-born细分方法。仪表和结果对不同参数影响紫外线辐射在地球表面。最后,太阳紫外线辐射的趋势进行了分析。从长期观察臭氧变化,由此产生的紫外线辐射的变化进行了讨论。未来预测水平的紫外线辐射是指在短时间尺度(天)以及可能的场景在未来几十年。
2.2理论
2.2.1能源来自太阳
天然紫外线辐射是太阳内部的核反应。产生的能量以这种方式向外传播通过太阳的气体身体一层称为光球层。光球层是层发出的辐射在地球上我们收到。它发出大约像黑体在5800 K,这是它的发射是连续在电磁波谱和光谱的形状是由普朗克定律决定的。光球层的温度是排放涵盖了从伽马射线光谱区附近的红外线(大约4000海里)(图1)。最大发射波长是由维恩定律
和太阳的温度这是0.5 /米(500海里),蓝绿色的可见光谱的一部分。然而,普朗克的形状曲线,人眼的相对灵敏度,可怕地依赖辐射与大气的相互作用(下面讨论)导致黄色太阳,我们观察。
太阳放射出的太阳辐射的总量是由斯蒂芬玻尔兹曼定律决定的
其中一个是斯蒂芬玻尔兹曼常数5.67 x 10 ~ 8 W m ~ 2 K-4。然而,入射能量在一个发光物体的距离与距离的平方根成正比,因此,能量到达地球大气层的顶端
卢比/平方= ctT4 r02
所以在哪里太阳常数,rs是太阳半径,r0平均日地距离。太阳能“常数”,最好的估计为1370 W m ~ 2,在多个时间尺度不同。对其温度、太阳的一生,因此其排放总量和光谱特性,改变了(据估计,排放增加了4.5 x 109年20 - 40%)。我们可以在时间范围内
T-I-i-I-i-I-i - | - | - | -i-i-r

图1所示。黑体的光谱在6000 K,太阳大气层外,太阳在地球的表面。
波长(nm)
图1所示。黑体的光谱在6000 K,太阳大气层外,太阳在地球的表面。
理解,太阳的活动,与可观测太阳黑子,22年的广泛的循环方式不同持续时间。然而,这包括太阳磁场的逆转和太阳黑子数目的周期(我们主要关心)11年。活跃的太阳黑子出现黑补丁的太阳和他们的磁场活动导致太阳耀斑——大爆发的能量增强紫外线和x射线辐射。这些,而不可预知的排放影响太阳常数,但在一个波长的依赖:峰变化波长160 nm的约为10%,而对于波长大于300纳米是小于1%,和太阳常数作为一个整体的0.1%。最后,太阳的27天旋转周期会导致几个百分比的变化在太阳紫外线输出,虽然在长波长(> 250海里)这种变化小于1%。
2.2.2行星运动和几何
短时间尺度变化,更直接相关的,是由于地球和太阳的天体运动。地球每年绕太阳稍微呈椭圆形,地球到太阳距离的变化,导致小的变化可用能源。当前的偏心轨道意味着地球离太阳(近日点)在1月南半球的夏季(北半球冬季)和离太阳最远的(远日点)7月,北半球的夏天。日地距离在两个极端之间的差异大约是3.4%,给不同的外星辐射约为6.9%。偏心本身在110000年周期变化(越来越少,然后椭圆),与极端的立场,给予不超过0.17%的地球入射通量的变化。近日点的位置也改变引力相互作用的结果(主要是与木星),导致地球的椭圆轨道进动,进而导致旋进春分和秋分的时机。随着时间的推移这些变化发生的时期18 800年和000年23年。时不影响的总能量受地球影响能量的方式分布在地球的表面。
最明显的变化太阳能得到在给定的位置、季节性效应引起的倾斜地球的轴。这个倾斜,地球的轴之间的夹角和黄道面,目前23.5°(22和24.5°之间的变化在一段时间内大约40000年)。它会影响白天的长度和高度的太阳在天空中,随时间变化和地球表面的位置。太阳是6月开销在北回归线(23.5°N),在春分和秋分(9月3月和)开销赤道,和12月达到了另一个极端位置在南回归线开销(23.5°S)。自地轴倾斜有微分阴影的纬度的乐队与头顶的太阳的位置变化(图2)和日长约等于无遮蔽的纬度圈的一部分。在春分和秋分这到处都是12个小时,而极地圈从24小时24小时黑暗的冬季日光在各自的夏天。天空中太阳的高度通常是表示太阳天顶角、z。这是当地垂直之间的角度和太阳的位置。太阳天顶角由因为给出z =罪9罪5 - f因为6因为年代因为h 6是纬度,5是太阳赤纬(纬度太阳6是纬度,5是太阳赤纬(纬度太阳在哪里
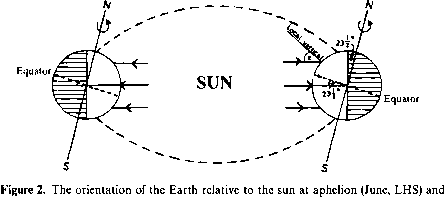
近日点(RHS) 12月。注意,太阳是分别为23.50°N和S开销,导致极地体验24小时光或黑暗的因为地轴的倾斜。
近日点(RHS) 12月。注意,太阳是分别为23.50°N和S开销,导致极地体验24小时光或黑暗,因为地轴的倾斜。
开销),中午和h时角。中午在当地太阳时角为零,增加15°(n / 12)每小时从中午。注意当地的太阳中午是经度和的函数不一定是一致的与当地时区(时钟时间)。当地太阳能中午进一步修改的“时差”,使年内变化约±15分钟,由于椭圆地球绕太阳和地球的倾斜的轴相对于飞机的轨道。日出和日落的因为z = 0时,导致的昼长一半的表达式H(因为H = -谭谭0 d
的能量水平表面的顶部上面的大气给定位置
Eo = S0 (r0 / r) 2因为z r是瞬时日地距离,并r0其平均值。
随着它进入大气辐射与大气成分的相互作用。大气中密度的变化、成分和温度的函数高度互动的类型和波长的辐射影响也高度的函数。表面我们观察的净效应在大气的深度衰减。
2.2.3大气
大气不是同质媒介。最多可以视为一系列统一的水平层,简化计算时,通常是通过大气辐射传输。在现实中许多大气特性可以改变在一系列空间和时间尺度。然而,物理可以讨论一维水平层的氛围。
垂直温度和密度大气的结构如图3所示,而低层大气的成分如表1所示。数密度,n(单位体积的气体分子数量)可以从理想气体定律确定n = P / kT次方
P是压力,T是绝对温度和k是玻尔兹曼常数(1.381 x 10-23 J K_1)。在大气压力和数量密度降低高度在一个近似指数(h)。恒温的假设下,应用的压力
H是一个规模高度约8公里低吗地区的大气。注意,n还取决于温度(见上图)既不是一个常数也不是一个简单的函数的高度对整个氛围(图3),所以n没有纯粹的指数与高度降低。
温度(K) 160 200 240 260
温度(K) 160 200 240 260
我miiiil我nii mal痛快哭。我问t迷你iliwil im
I0 1 ~ 3“,”来自我»0 10 * I03
图3。垂直压力,密度和温度标准大气。表1。低的组成大气的一部分(没有水蒸汽)
我miiiil我nii mal痛快哭。我问t迷你iliwil im
I0 ~ 3
这篇文章有用吗?
推荐项目


读者的问题
-
艾嘉Pennanen1个月前
- 回复
-
安东尼奥1个月前
- 回复
-
吉拉尔多2个月前
- 回复
-
Elisa2个月前
- 回复
-
andwise1年前
- 回复
-
斯蒂芬妮1年前
- 回复
-
斯蒂芬妮1年前
- 回复
-
哈利1年前
- 回复
-
尽管阿兰1年前
- 回复
-
茉莉花took-took1年前
- 回复
-
1年前
- 回复