新墨西哥州中南部数十年干旱周期的模式和后果
布鲁斯·t·米尔恩·道格拉斯·i·摩尔胡利奥·l·贝当古·詹姆斯·a·帕克斯·托马斯·w·斯威特南·罗伯特·r·帕门特·威廉·t·波克曼
极端区域干旱是半干旱生态系统中最常见的干扰形式,其典型特征是恢复速度相对较慢。干旱导致的影响包括区域同步的昆虫爆发、野火和树木死亡(Swetnam和Betancourt 1990),以及农业、造林和畜牧业的灾难性失败(Mainguet 1994)。干旱条件,伴随着人为的土地管理不善,导致了随后的入侵草原和农田木本灌木以及非本地的蛙类和草类,导致了在世界许多地方表现出来的现代“沙漠化”过程(Archer et al. 1988)。
在美国西南航空在美国,20世纪50年代的干旱是过去千年中最严重的气候事件之一,因为它对该地区的生态产生了广泛的影响(Herraybet雷竞技最新bel et al. 1972;Swetnam和Betancourt 1998),水资源(Thomas 1963)和经济(Regensberg 1996)。随着西南地区人口和资源需求的增加,经济对大规模干旱的敏感性也会增加。清楚地了解极端干旱不仅是了解长期生态系统动态的必要条件,也是减轻社会经济影响的必要条件。
本章的目标是利用新墨西哥州中部的塞维利亚LTER站点(1)量化从394年树木年轮记录推断的降水的年代际变化,(2)将降水的重复年代际波动与19世纪90年代和50年代的重大干旱联系起来,(3)评估与上世纪干旱相关的生态响应,以及(4)阐明可能影响未来响应的生物-大气反馈。我们评估了年降水的年代际波动的量级、时间和后果。
塞维利亚塔LTER站点描述
塞维利亚LTER研究基地位于新墨西哥州索科罗县(34°20′N, 106°50′W)的塞维利亚国家野生动物保护区(NWR)。塞维利亚国家野生动物保护区包括10万公顷的草地、沙漠和林地山脉以及中间的格兰德山谷。海拔范围从里约热内卢Grande的1350米到保护区西北部Ladrón峰的2797米。地形、地质、土壤和水文,与主要的气团动力学相互作用,提供了一个空间和时间模板,使该地区成为几个生物群落之间的过渡区。该地区包括代表大平原草原、大盆地灌丛草原、奇瓦瓦沙漠、内陆灌木丛和山地针叶林的群落(Brown 1982)。
塞维利亚气候描述raybet雷竞技最新
塞维利亚LTER研究区域横跨主要季节气团之间的边界(例如,冬季北极锋面系统向南穿越大平原,影响塞维利亚东部边缘;大盆地极地气团延伸到塞维利亚北部边缘;百慕大高地在山上形成夏季对流风暴,沿着塞维利亚的低地向东北移动)。叠加在这些空间模式上的是El Niño-Southern振荡(ENSO)现象的时间动态。这些气候现象raybet雷竞技最新进一步转化为地形南落基山脉.
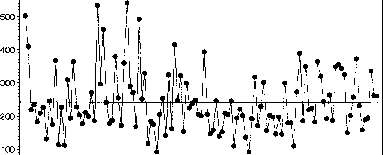
塞维利亚塔地区的年度气raybet雷竞技最新候包括两种不同的季节性风暴模式。在晚秋、冬季和春季,风暴会在东北部和热带太平洋形成。这些风暴是由极地和亚热带引导进入该地区的急流,它们的年频率和新墨西哥州中部的总降水量在一定程度上受到年际(ENSO)和年代际(太平洋年代际振荡)太平洋气候的变化。raybet雷竞技最新一般来说,新墨西哥州中部在El Niño事件期间,或太平洋年代际振荡(PDO)为正值时,往往有湿润的秋季、冬季和春季,而在La Niña事件期间则相反(ENSO,见Andrade和Sellers 1988年,Molles和Dahm 1990年,Redmond和Koch 1991年,Cayan和Webb 1992年,Kahya和Dracup 1993年;PDO参见Cayan et al. 1998, Mantua and Hare 2002)。在塞维利亚塔以南的索科罗(Socorro),与过去80年的中间年份相比,El Niño年10月至5月期间的降水量增加了53%,La Niña事件期间的降水量减少了略超过一半(图15.1,表15.1)。虽然这种秋冬春季降水的增加可能看起来很小,但较冷的条件和休眠的植被有利于土壤水分的补充,从而增加了生长季节土壤水分的有效性。此外,大多数这些凉爽季节的锋面风暴都有宽阔的轨迹
降水毫米600:
我的1 1 1 1 1“我”“我1”1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1的1我' 1 ' 1 1 1 1 1 1 1 1 1
1860 1890 1900 1910 1920 1933 1940 1950 1930 1970 1980 1990 2000
一年
我的1 1 1 1 1“我”“我1”1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1的1我' 1 ' 1 1 1 1 1 1 1 1 1
1860 1890 1900 1910 1920 1933 1940 1950 1930 1970 1980 1990 2000
一年
图15.1新墨西哥州索科罗的降水记录(1880-2000年)。注释周期低降水从19世纪90年代末到20世纪初,以及1950-1957年。
(>100公里),并倾向于同步整个区域的生态系统对降水事件的响应。
夏季降水是由北美季风驱动的。在夏季,急流向更北纬的地区退却,从而减少了太平洋风暴系统对该地区的影响。通常情况下,西南地区最大的降水出现在7月、8月和9月,并与北美季风期间的对流雷暴有关。季风降水源于墨西哥湾上空的潮湿气团,并被百慕大高压推向西南(Mitchell 1976;尼尔森1986)。由此产生的降水由雷暴在地形上不均匀地分布,雷暴起源于山脉,移动到低地。虽然时间变异性低,但空间变异性高
表15.1 1914-1993年新墨西哥州索科罗按ENSO等级划分的平均降水量
降水(毫米)
ENSO
年度10月- 5月- 6月
El Niño 15 275.8 a 156.2 a 119.6 a
内侧56 239.4 a 102.3 b 137.1 a
La Niña 9 162.5 b 49.9 c 112.5 a
a不同字母的ENSO分级差异有统计学意义(P < 0.05)。
高。ENSO现象与新墨西哥州夏季降水之间的联系较弱(Andrade and Sellers 1988;Molles等,1992;Harrington et al. 1992)。对于季风湿度的开始和/或强度似乎有一定的可预测性(Higgins和Shi 2000),但可预测性远远低于与ENSO周期极端相关联的冬季和春季湿度。虽然7月、8月和9月的季风水分通常占塞维利亚地区年降水量的一半以上,但高蒸散量阻止了夏季水分将土壤水分补充到深处。因此,植被对夏季水分的反应是短暂的和零星的。
因此,我们认为秋季-冬季-春季降水与夏季(6 - 9月)降水是分开的。对塞维利亚土壤湿度的持续监测表明,5月和6月气温的升高和植物的生长消耗了之前非季风期的表层土壤水分季风七月开始。季风的水分通常在两次风暴之间通过蒸发迅速流失(2-3天),因此从夏季到冬季的过渡时期通常也是土壤干燥的时期。
多年代际降水模式
塞维利亚LTER的直接降水测量始于1988年,而索科罗县的记录仅可追溯到19世纪70年代。因此,我们从树木年轮获得的年降水记录中阐明了持续的年代际天气模式。帕克斯等人(已出版)通过结合塞维利亚NWR东段洛斯皮诺斯山脉阿罗约·德·米拉格罗地区的活树和死树(因20世纪50年代干旱而死亡)的皮松(Pinus edulis)年轮年表,重建了394年的降水历史(1598-1991年)(图15.2,15.a)。原始重建降水深度表示为与长期平均值(286 mm, s.d, = 63.5)的偏差,以强调异常的干湿年份(图15.a)。这种特殊重建的细节在Betancourt等人(1993)和Parks等人(出版中)中有解释。塞维利亚树木年轮指数年代学与索科罗重叠时期(1892-1991年)实测的实际降水量进行了统计比较。我们比较了几个季节,但相关性最高(r = 0.74)的是13个月的间隔,涵盖了生长年度的前8月到当前8月。对于新墨西哥州南部的针叶树,Grissino-Mayer等人(1997)发现了类似的形成层生长受到前一个夏秋季节和当前春夏季节降雨的制约,冬季降雨(1月至3月)影响不大。利用树轮指数对8 - 8月降水量进行线性回归。
我们采用多锥谱分析方法(MTM;Dettinger et al. 1995)来测量重建降水记录的时间变异性。MTM频谱分析使用三种正交波形来测量时间序列中不同频率或相反周期的方差。置信区间(95%和99%)是在地球物理适当的零假设下定义的
-

- 图15.2新墨西哥州塞维利亚塔国家野生动物保护区洛斯皮诺斯山脉的松柏林地。该山区的皮松为重建长期降水记录提供了样本。

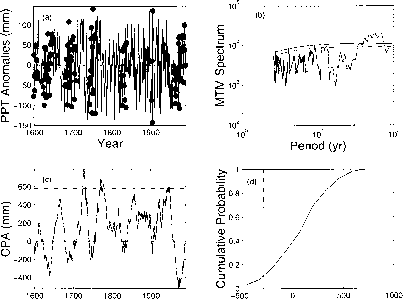
时间序列是红噪声,也就是说,比白噪声更相关,比随机游走更不相关。因此,MTM谱中超过置信限的峰值表明持续振荡(图15.3b)。在年代际尺度上,塞维利亚树轮重建在72.9年、68.5年、64.1年、60.2年、56.8年、53.8年、51.3年、48.8年、44.4年、42.7年、40.9年、39.4年和36.6年(平均= 51.9年,s.d = 11.3年)具有显著的(99%置信区间)变异性。因此,在阿罗约·德·米拉格罗平均每52年就会发生一次干旱和湿润期。必须谨慎地评估这些数据,因为在原始树年轮记录的构建过程中,去趋势可能导致对低频变化的高估(D. Meko, pers。通讯,2002)。
干旱给植物带来了相互矛盾的人口挑战。种子处于这样一种竞争中:(1)出现适合发芽的条件;(2)生存或逃脱捕食的机会不断减少。干旱会延缓前者,可能会加速后者。因此,对年降水量记录进行生态相关分析时,应关注与平均降水量的偏差,因为偏差衡量的是任何给定年份的唯一性。此外,一连串潮湿的年份有利于需要异常大量水分的物种的萌发和建立,如蓝色的格鲁马草,布特拉·格鲁拉(Neilson 1986)。一连串的干旱年份将耗尽种子库,从而使植物群落在未来几年的动态变得丰富多彩。因此,我们将降水量的年度估计重估为偏离平均值或“正常值”(图15.a),并检查了连续偏差的字符串,以找到持续湿润和干燥条件的时代。
注册会计师(毫米)
图15.3基于Arroyo de Milagro树轮记录的降水长期变化格局。(a)年降水深度减去长期平均值。点表示发生在干旱时期的年份。(b)原始降水深度的多锥法谱。水平线表示95%(虚线)和99%(实线)置信区间。(c)降水异常的运行和,即累积降水异常CPA。(d)注册会计师的累积概率分布。
注册会计师(毫米)
图15.3基于Arroyo de Milagro树轮记录的降水长期变化格局。(a)年降水深度减去长期平均值。点表示发生在干旱时期的年份。(b)原始降水深度的多锥法谱。水平线表示95%(虚线)和99%(实线)置信区间。(c)降水异常的运行和,即累积降水异常CPA。(d)注册会计师的累积概率分布。
继Feller(1968)和水文学中的“质量曲线”技术(例如,Peng和Buras 2000)之后,我们通过形成异常的运行总和,即累积降水异常(CPAs),揭示了持续的、重复的干湿期(图15.3c)。峰值和低谷分别对应净累积量的高低。最大CPA (787 mm)出现在1727年,最低CPA (-523 mm)出现在1971年。生活在1727年或1971年的植物分别处于极湿或极干时期的末期。CPA的累积概率密度表明,只有10%的年份处于17世纪早期和50年代这样的严重干旱(图15.3)。相反,异常的倾斜分布产生的湿期在20%的时间内超过了CPA的一个标准偏差。个别年份在某种程度上更有可能处于湿润时期而不是干燥时期。
有7个长串的干旱年份构成了持续干旱。将记录长度除以7表明56年是干旱重现的大致时间间隔,这与时间序列分析一致(图15.3b)。这段时间间隔与19世纪90年代和50年代的大旱相吻合,两次大旱相隔约60年。
我们的结论是,降水的长期波动发生在
塞维利亚地区大约每52年进行一次,尽管41年至63年的间隔与预期52年的间隔在一个标准偏差内。预测未来干旱的尝试受到复发间隔的巨大变化和缺乏关于振荡的气候起源的确定理论的限制。然而,这一限制并不否认过去存在过长达十年之久的干旱,这种干旱可能会再次发生,造成重大的生态和社会经济后果。
年代际至多年代际气候振荡的可能来源raybet雷竞技最新
塞维利亚树轮系列低频变化的最终来源目前尚未解决。年气温和降水序列在年代际到世纪尺度上的振荡模式已被确定仪器记录(例如,Cayan et al. 1998;麦凯布和德廷格1999;Dettinger et al. 2001)和北美西部的树木年轮(例如,Biondi et al. 2001;Gedalof和Smith 2001;Villalba et al. 2001)。据推测,这种低频变化大部分源自太平洋盆地,并涉及ENSO模式与较长期的十年至百年气候波动的相互作用(Dettinger et al. 2001)。raybet雷竞技最新注意大西洋也发现了类似的低频变化(例如,Delworth等人,1993年;Enfield and Mestas-Nunez 1999),并能产生相似的温度和降水周期,特别是在夏季。在北太平洋,大部分海面温度变化以年代际(~20-30年)时间尺度的模式发生,并伴随着冬季阿留申低压强度和位置的变化。这就是所谓的北方太平洋振荡(禁食;Gershnov and Barnett 1998)或太平洋年代际涛动(PDO;曼图亚和野兔2002)。
对诸如PDO这样的低频变化的主要解释是随机大气强迫、大气遥相关、中纬度海洋-大气相互作用、热带-热带外相互作用、海洋遥相关和内在海洋变率(见Hare et al. 2000的摘要;Dettinger等,2001;曼图亚和野兔2002)。关于稳态和混乱的行为十年到世纪尺度的变化,以及它的可预测性。乐观的观点是,关于长期模式的当前阶段的知识(例如,PDO)可以用于预测未来几年的气候(Latif和Barnett 1996;raybet雷竞技最新Dettinger等人,2000;Schneider and Miller 2001)。
20世纪有两个完整的PDO周期。1890年至1924年和1947年至1976年,“冷”或负PDO(更像拉尼娜)政权盛行,而“暖”或正PDO政权盛行于1925年至1946年和1977年至1994年。Minobe(1997,1999)利用小波分析发现,北太平洋海面温度、海平面压力和基于北美树木年轮的温度重建的波动在15-25年(北方冬季)和50-70年(北方冬季和春季)的周期中最为活跃。根据Minobe(1999)的说法,这两个周期与三个周期的相对周期同步,并在20世纪20年代、40年代和70年代当它们反转时在北太平洋气候中产生了“政权转移”。raybet雷竞技最新从1998年开始,有轻微的迹象表明这种模式的转变,当时PDO指数急剧转为负值(“凉爽”模式),热带太平洋在1976年之后的长时间变暖后开始变冷,北大西洋在20世纪60年代以来的长时间变冷后开始变暖。这就是20世纪40年代政权更迭期间的情况,导致了20世纪50年代的干旱,根据塞维利亚的树木年轮,这种情况往往每41-63年发生一次。我们抵制诱惑,不做任何长期预测,但我们建议,新墨西哥州的政治家、资源经理和牧场主都没有什么理由乐观。正如Swetnam和Betancourt(1998)所指出的,考虑到另一场极端干旱的可能性,当地生物学家应该准备好利用这种“自然实验”。
Arroyo de Milagro重建用于区域评估的可靠性
我们的分析的年轮降水年代学表明,塞维利亚的Arroyo de Milagro每41-63年发生一次大干旱(图15.3b,c)。树木年轮记录与现代气象测量之间的重叠使我们能够评估塞维利亚记录在多大程度上反映了更广泛的中格兰德盆地的情况。
测量的年总降水量不能很好地预测生态系统生产力因为降水的季节时间对其生物有效性至关重要。时间和土壤持水能力等其他因素会影响降水对生长的影响(Valentine and Norris 1964)。帕尔默干旱严重指数(PDSI;Palmer 1965)可以识别生态有效的干湿条件。PDSI使用降水、温度和土壤湿度来衡量水分有效性。该指数是根据当地气候进行标准化的,因此可以比较整个美国的相对干燥和湿润程度。raybet雷竞技最新PDSI是外推的raybet雷竞技最新气候地区在州内。
塞维利亚地区的PDSI记录涵盖了从1895年到现在的整个时期(图15.4;源http://lwf.ncdc.noaa.gov/oa/raybet雷竞技最新climate/onlineprod/干旱/ main.html)。指数高于0的时期被认为是湿润时期,而低于0的时期则是干燥时期。干旱分类从-2.0开始,随着指标值的降低而加重。在塞维利亚LTER相对较短的时期内(1989-至今),情况从1989年的干旱La Niña到1992-1993年El Niño期间的相当潮湿。此后,该指数基本保持干燥状态,直到1996年的湿润季风,该季风持续到1997-1998年的厄尔Niño,然后在1998年延长的拉Niña,直到现在(2002年),基本上保持干燥状态。从湿到干的振荡显然是常态。
这种模式最明显的例外发生在19世纪90年代至20世纪初和1950年至1957年这两个较长的时期。此后,该指数一直保持在零以下。19世纪90年代到20世纪初的干旱是罪魁祸首
帕尔默干旱严重指数
1979 T T 1984 ^ T 1SSST r T 1994 20 02
帕尔默干旱严重指数
1979 T T 1984 ^ T 1SSST r T 1994 20 02
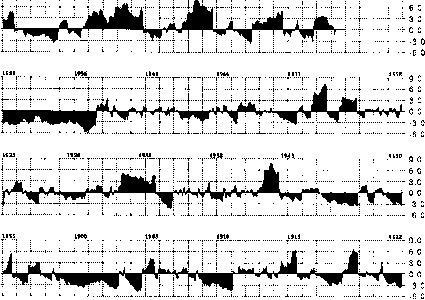
图15.4新墨西哥州中部帕尔默干旱严重程度指数(1895-2002)。请注意1898-1905年和1950-1957年期间的低(干)指数时期。
新墨西哥州-第05组:189S-2002(每月平均)
图15.4新墨西哥州中部帕尔默干旱严重程度指数(1895-2002)。请注意1898-1905年和1950-1957年期间的低(干)指数时期。
19世纪90年代,西南牧场大量牲畜死亡和灌木入侵。1950-1957年这段时期被称为“50年代干旱”。在西南部,这比20世纪30年代俄克拉何马州和德克萨斯州狭长地带的“沙尘暴”干旱要严重得多。表15.2列出了年PDSI平均值小于- 2.0的年份。PDSI所指示的这两个干旱期与树木年轮年表并反映低降雨量索科罗降水记录的时期。
PDSI值与重建降水深度显著相关。利用树木年轮重建降水的回归分析(Betancourt et al. 1993;Parks et al.出版)和气候区5 (Middle里约热内卢Grande Valley)的年平均PDSI值得出R2raybet雷竞技最新 = 0.46 (p < 0.0001, n = 95;图15.5)。其中,5月和6月相关性最强(R2 = 0.497, p < 0.0001, n = 95, R2 = 0.469, p < 0.001, n = 95)。因此,重建的降水深度与区域水分条件有统计学相关性。然而,由于Arroyo de Milagro遗址的土壤、水力和地形条件,这种无法解释的巨大变化提醒人们不要将该记录肆无忌惮地应用于该地区。
一年 |
PDSI |
一年 |
PDSI |
一年 |
PDSI |
一年 |
PDSI |
1900 |
-3.12 |
1918 |
-2.59 |
1946 |
-2.50 |
1971 |
-2.25 |
1901 |
-2.44 |
1922 |
-2.41 |
1947 |
-3.01 |
1982 |
-2.27 |
1902 |
-2.65 |
1934 |
-2.56 |
1950 |
-2.09 |
||
1904 |
-3.29 |
1951 |
-3.38 |
||||
1909 |
-2.39 |
1952 |
-3.02 |
||||
1910 |
-3.19 |
1953 |
-2.30 |
||||
1954 |
-3.31 |
||||||
1955 |
-3.11 |
||||||
1956 |
-3.78 |
||||||
1957 |
-3.23 |
气候对塞维利亚的生物群落过raybet雷竞技最新渡区的影响
塞维利亚LTER站点位于几个站点的接口主要生物群落包括短草草原、奇瓦汉沙漠灌丛和松柏林地。潜在蒸散量和有效土壤水分的综合作用似乎调节了土壤水分的分布各种各样的生物群落.例如,当均匀的土壤出现在局部粗糙的地形上时,坡度和坡向的细微差别调节了地表能量供应,并产生重复的植被模式(图15.6)。沙漠灌木,如三叉戟Larrea占主导地位朝南的斜坡高水分亏缺。几米远的地方,在同一海拔高度上,有一种叫Juniperus monospma的植物北坡.对20世纪50年代干旱前后拍摄的航拍照片的检查表明,在干旱期间条件变得贫瘠的东部和西部斜坡上发现的杜松死亡。今天,这两种优势物种在这样的地区相遇。重复摄影显示,L. tri-dentata在20世纪50年代的干旱之后变得更加广泛(图15.7),显然反映了新墨西哥州沙漠灌木丛的大规模扩张(Grover和Musick 1990)。
我们初步测量了L. tridentata和J. monosperma的相对生长速率作为年降水量的函数,以检验该物种在遥远的过去能够在Arroyo de Milagro生长的假设。在过去400年里,周期性的干湿交替(图15.3c)应该为L. tridentata等物种的建立创造了机会。然而,令人满意的资源可得性并不能克服分散限制或竞争,这些限制或竞争也会影响建立。正在进行的现场实验解决了有竞争力的相互作用这些物种的。
在7个地点(200 三叶松生物量(B)变化为dB/Bdt = -137.2 + 1.28 PPT - 0.0029 PPT2,其中PPT为2000年年降水量(mm)。对于单精子J. dB/Bdt = -6.83 + 0.05 PPT-0.0001 PPT2。我们模拟年净初级生产力在1598-1991年间,假设植物开始时的生物量为1公斤/平方米(即不受扩散的限制),并有潜力以后勤方式增长到50公斤/平方米的承载能力。我们假设没有竞争。因此,根据树木年轮记录估计的年降水深度,模拟估计了每个物种的潜在净初级生产力。 Arroyo de Milagro相对较高的年平均降水量更有利于单精草的生长。几十年来,刺柏产量持续处于高水平,同时也出现了L. tridentata的致命条件(图15.8)。适宜生长的条件在整个记录中都有出现,但被不生长的时期反复打断。我们认为,气候变化、扩散限制和竞争可能是导致该地区直到最近才出现三叉叶蝉的原因。 当然,更大范围内的变异性可以维持两个物种的源种群。源种群可以通过注入繁殖体来改善特定地点气候变化的影响。raybet雷竞技最新因此,一个全面的观点将包括一个元人口的方法来理解的作用时态变化通过空间(Keymer et al. 2000)。raybet雷竞技最新气候波动会改变种群的边缘,因为物种会超越它们以前的分布范围,或者从被占领的地区被消灭。类似地,在一个物种分布的边缘,小气候的变化可能会造成局部的斑块分布(Holt和Keitt 2000)。raybet雷竞技最新 与干旱生态系统特别相关的生理极限的一个例子来自于植物维持蒸腾作用对水的需求。通过木质部的水运输受到空化的干扰,空化是由干旱引起的高木质部张力或干旱周期引起的冻结和解冻在木质部(Tyree和Sperry 1989)。引起空化的干旱或冰冻条件在不同物种之间差异很大,由木质部的结构特征、干旱情况下的坑膜孔径(Sperry和Tyree 1988)和冰冻情况下的木质部导管体积(Davis et al. 1999;Sperry et al. 1994)。这种与木质部结构的可量化联系使得植物功能物理极限的定义比一些功能极限难以预测的生理参数更直接。虽然一些物种可以修复空化的影响(Holbrook和Zwieniecki 1999),但个体的水力极限在很大程度上是固定的,使得个体的水运输维持依赖于它们在活动时经历的干旱和冰冻条件。 高度可变的降水减少了有利于发芽和建立的土壤水分条件以必要顺序出现的机会(Neilson 1986)。虽然塞维利亚森林的优势木本树种(例如,edulis Pinus, J. monosperma, L. tridentata)具有高度耐旱性(Linton et al. 1998;痘痕, man和Sperry 2000),有限的幼苗根系暴露在极干燥的浅层土壤中。在这种条件下,大量的空化会导致死亡(Williams et al. 1997)。一旦建立,优势灌木物种只有很少接近完全空化的点(Pockman和Sperry 2000)。长期干旱时期的条件,如50年代新墨西哥州的干旱(Allen和Breshears 1998年),很可能接近这些物种对干旱引起的空化的生理极限。 在塞维利亚,从以L. tridentata为主的灌丛向北部的草地过渡,是L. tridentata连续分布的北部界限。虽然它的常绿习性使它能够在任何时候利用有利的条件,但它也必须在冰冻条件下保持水分运输,以支持它的常绿叶子。在其北部极限的L. tridentata的历史报告表明,极端冰冻事件导致地上生长的严重枯死(Cottam 1937)。在亚利桑那州南部,成熟的三叉戟(L. tridentata)茎在0°到-10°C之间冻结后没有栓塞,之后栓塞率随温度的升高呈线性增加降低温度直到-16°至-20°C完全栓塞(Pockman和Sperry, 1997年)。对长期气候数据的粗略分析表明,这一临界温度范围与索诺拉沙漠和莫哈韦沙漠的物raybet雷竞技最新种北部极限相对应,但不能解释格兰德山谷的范围向北延伸到塞维利亚的极限。最近的数据表明,塞维利亚的L. tridentata比亚利桑那州的L. tridentata更能抵抗冰冻引起的空化(Martinez-Vilalta和Pockman, 2002)。生物地理变异提供了一个机会来检查这一生理极限在决定物种分布中的作用。 塞维利亚气候的生态系统反馈raybet雷竞技最新 虽然温度和湿度明显影响塞维利亚LTER遗址的植物分布,但有一些证据表明,植被模式也对当地气候有反馈。raybet雷竞技最新其中一个例子发生在塞维利亚的一个地区,该地区以L. tridentata为主(图15.7)。人们普遍假设,墨西哥和美国西南部干旱地区的人为干扰与干旱周期一致(例如,19世纪90年代和50年代的大旱;黑斯廷斯和特纳1965),促进了木本灌木的范围扩大的最近加速。在19世纪后期,牧场被过量的牛、羊和马过度放牧,加上持续的干旱,偏爱的沙漠灌木(L. tridentata和牧豆树,豇豆spp.)。Bray (1901, p. 289)指出了世纪之交入侵西德克萨斯州的速度:“关于木本植被的建立,长期观察的人们一致认为,大多数灌木[=L。和豆科植物覆盖的地区以前是开阔的草原。”二十世纪,沙漠灌木继续向北侵入草原,特别是在新墨西哥州和西德克萨斯州(Gardner 1951;Branscomb 1958;汉弗莱1958; Buffington and Herbel 1965; York and Dick-Peddie 1969; Hen-nessy et al. 1983; Humphrey 1987; McPherson et al. 1988; Grover and Musick 1990). 灌丛物种入侵草原不仅对生态系统动态(Schlesinger et al. 1990)和人类活动(Mainguet 1994)产生重大影响,而且还可能改变中尺度气候条件。Hayden(1998)回顾了现有文献,分析了大尺度温度模式,并与观测到的最低温度相关在沙漠美国西南部的露点温度与相同地点的预测露点温度相差很大。冬季最低气温比预测的高8°C (Hayden 1998年);在塞维利亚LTER站点,冬季最低温度可能比露点温度预测的高4°C。造成这种差异的原因是由于从大气中释放出的非甲烷碳氢化合物沙漠植被(特别是来自沙漠灌木如L. tridentata的萜烯);这些碳氢化合物起到了“温室气体”的作用,降低了植被上方当地大气的辐射率,防止了热量损失,提高了夜间最低温度。 在塞维利亚LTER站点,两个气象站位于同一海拔麦肯齐平原上,相距不到5公里;一处为三叉戟占优势的群落,另一处为草地所包围。在木酚灌木植被的站点通常记录较高的夜间最低温度 温度°C 25 20 15 25 20 15 1 23456789 一年中的一天站40 =草地49 =灌木40 1 23456789 一年中的一天站40 =草地49 =灌木40 图15.9塞维利亚LTER站点2000年1月1日至10日沙漠灌木植被和草地植被上空气温;气象站相距4.4公里。注意沙漠灌木植被上较温暖的最低温度。 (图15.9),特别是在风平浪静期间(大风混合了两个站点的大气)。大气变暖降低了霜冻死亡的概率,从而增加了这些沙漠灌木物种的生存可能性和种群数量的增加。因此,这可能是由于灌木的入侵加速了草原的沙漠化,通过大气化学的变化,最终改变了温度制度,通过减少霜冻,可以提高灌木种群的存活率和数量。未来需要对这一假设进行研究,以确定植被变化是否以及在多大程度上影响干旱和半干旱生态系统的区域气候。raybet雷竞技最新 生态学家在测量或模拟生物对气候较长周期的反应方面进展缓慢。raybet雷竞技最新例如,大多数间隙模型将气候变率模拟为围绕年平均值的随机方差。raybet雷竞技最新然而,在间隙模型中插入年代际气候波动已被证明要么诱导周期性,要么改变生物量的平均值(Yeakley et al. 1994)。塞维尔莱塔的一般特征raybet雷竞技最新气候包括季节性、年际和年代际变化的尺度。 年降水量的41- 63年周期构成了一个缓慢变化的气候背景,在这个气候背景下,发生了更高频率的生态响应。管理、疾病或捕食对野生种群的改变可能导致不同的反应,这取决于振荡是上升还是下降。重新造林或土地开垦的努力可能在周期的湿润阶段最为成功,在此期间20至30年的相对潮湿的天气将有利于这些努力。在随后的振荡干燥期维持牧场边际生产的尝试有失败的风险。事实上,塞维利亚国家野生动物保护区是在20世纪50年代干旱结束后仅10年,即CPA处于历史最低水平(图15.3c),从一个工作的牧场建立起来的,这可能并非偶然。 政策制定者和金融机构可能会预见到这一点经济崩溃随之而来的干旱刺激了土地所有权的突然变化。塞维利亚的树木年轮记录只记录了7次不同严重程度和持续时间的多年干旱,表明重大干旱的重复周期为52年(图153亿),最后一次干旱持续在1942年至1972年之间。我们目前缺乏必要的时间深度、区域覆盖范围,以及对构成这种明显周期性的全球尺度海洋-大气相互作用的理解。我们还认识到,这些周期性可能是固有的不稳定的,并可能受到各种现象的调节,包括人为气候强迫。raybet雷竞技最新因此,我们在对新墨西哥州中南部即将发生的干旱做出任何预测时都要谨慎。尽管如此,我们指出,21世纪初的极端干旱是可以预料的,新墨西哥州中南部的人口增长和对水和其他资源的需求扩大了对干旱的敏感性。 土地管理者可能会反思19世纪90年代的经历。铁路的引入刺激了人们储备这一品种,因为这样更容易进入市场。同时,风车的引入使牧场主得以供应饮用水在干旱时期饲养牲畜。然而,如果没有降水,草料植物无法承受高载畜率,导致土壤有机质和种子库长期耗竭。在未来的干旱中,明显的技术解决方案如精确耕作方式就像过去的水井和火车一样,优化的灌溉策略和转基因作物或牲畜实际上可能会增加景观或生态系统各个组成部分的风险。半干旱土地的生态受到人类职业生涯顺序的环境变化的影响,因此可能应该在人类几代人的时间尺度上进行管理。 的变化降水发生在季节、年际和十年的时间尺度上。与土地使用和资源管理有关的生态和经济后果促使人们从直接和替代气候记录中寻找可重复的模式。raybet雷竞技最新我们研究了来自塞维利亚塔国家野生动物保护区piñon松(Pinus edulis)年轮的394年(1598-1991)降水记录 美国新墨西哥州中部的避难所和长期生态研究地点。19世纪90年代和50年代的主要区域干旱与降水量的52年显著周期性(标准差为11.3年)一致。十年干旱的长期生态后果涉及新物种的建立、生理压力、植物与大气之间的反馈以及与土地使用有关的经济影响。模拟的单籽刺柏(Juniperus monosperma)和油木灌木(Larrea tridentata)的净初级生产力表明,在过去的四个世纪里,适合刺柏生长的条件断断续续地出现。对19世纪90年代和50年代干旱的评估表明,未来的技术尝试将改善干旱干旱的影响应尽量减少景观的各个组成部分的不可预见的后果。年降水量偶然的年代际振荡是该地区的一个主要生态因素。 Dave Meko, Mike Dettinger和两位匿名评审员指导了这项工作的最终结果。NSF为BTM提供了DEB 9910123和DEB 0080529的资助。塞维利亚LTER出版号。263. 艾伦,C.和D.布雷希尔,1998。干旱引起的森林-林地过渡带的转移:对气候变化的快速景观响应。raybet雷竞技最新美国国家科学院论文集95:14839-14842。 安德拉德,e.r.和W.D.塞勒斯,1988年。El Niño及其对亚利桑那州降水的影响。气候杂志8:403-410。 阿彻,S. C.斯克利夫雷斯,C.R.巴沙姆和R.马乔,1988。亚热带热带稀树草原的自生演替:草地向刺林的转变。生态专论58:111-127。雷竞技csgo 贝当古,j.l., E.A.皮尔逊,K.A.赖兰德,J.A.帕克斯和J.S.迪恩。1993。历史和气候对新墨西哥州松柏林地的影响。raybet雷竞技最新为可持续发展和社会需求管理杜松生态系统第53-62页。美国农业部林业局通用技术报告RM-236。落基山森林和山脉实验站。美国农业部。科罗拉多州柯林斯堡。 Biondi, F., A. Gershunov,和D.R. Cayan, 2001。自1661年以来北太平洋年代际气候变化。raybet雷竞技最新气候杂志14:5-10raybet雷竞技最新。 布兰斯科姆,b.l., 1958。新墨西哥南部沙漠草原范围的灌木入侵。牧场管理杂志11:29 -132。 布雷,1901年。德州西部植被的生态关系。植物学报32:99-123,195-217,262-291。 布朗,1982年。美国西南部的生物群落——美国和墨西哥。沙漠植物4:1-342。 Buffington, l.c.和C.H. Herbel, 1965。1858 - 1963年半荒漠草地植被变化。生态专论35:139-164。 卡扬,d.r.和R.H.韦伯,1992。El Niño/美国西部的南方涛动和气流。第29-68页在H.F. Diaz和V. Markgraf,编辑,El Niño南方涛动的历史和古气候方面。剑桥大学出版社,剑桥。 卡扬,d.r., M.D.德廷格,H.F.迪亚兹和N.E.格雷厄姆,1998。北美西部降水的年代际变化。气候杂志:3148-3raybet雷竞技最新166。 科塔姆,1937年。犹他州已经失去了索诺兰地区下游的所有权了吗?科学85:563 - 564。 戴维斯,S.斯佩里,U.哈克,1999。木质部导管直径与冰冻空化的关系。美国植物学杂志86:1367-1372。 德沃斯,T., S. Manabe,和R.J. Stouffer, 1993。耦合海洋中温盐环流的年代际变化大气模型.气候学报6:19932raybet雷竞技最新011。 戴廷杰,M.D hil, C.M. Strong, W. Weibel, P. Yiou, 1995。软件加速噪声时间序列的奇异谱分析,Eos, Trans。美国地球物理学会76(2):12,14,21。 德廷格,医学博士,D.R.卡扬,G.J.麦凯布,Jr.和K.T.雷德蒙,2000。2001年冬春美国基于预期中性ENSO条件和最近NPO状态的流概率。实验长程预报9:55-60。 德廷格,m.d., D.S. Battisti, R.D. Garreaud, G.J. McCabe和C.M. Bitz, 2001。年际和年代际enso样气候变化对美洲的半球间影响。raybet雷竞技最新V. Markgraf主编《美洲现在和过去的半球间气候联系及其社会影响》第1-16页。raybet雷竞技最新学术出版社,圣地亚哥,加利福尼亚。 恩菲尔德,d.b.和A.M.Mestas-Nunez。1999.全球海面温度的多尺度变化及其与对流层气候模式的关系。raybet雷竞技最新气候杂志:2719-2733。 Feller, 1968年。概率论及其应用简介。卷。1。威利,纽约。 加德纳,1951年。新墨西哥州格兰德山谷木酚灌木地区的植被。生态专论21:379-403。 盖达洛夫,Z.和D.J.史密斯,2001。太平洋北美的年代际气候变率raybet雷竞技最新和体制尺度变化。地球物理研究快报28:1515-1518。 Gerschnov, A.和T.P. Barnett, 1998。ENSO遥相关的年代际调制。美国气象学会通报79:2715-2725。 h.d., C.H.拜桑和T.W.斯威特南。1997。格兰德盆地南部1374年降水重建。最终报告,国防部,遗产计划,布利斯堡,德克萨斯州。 格罗弗,h.d.,和H.B. Musick, 1990。美国新墨西哥州南部灌木丛的侵蚀:美国西南部沙漠化过程的分析。气候变化:305-330。 海尔,s。r。s。米诺贝,w。s。伍斯特,s。麦金内尔。2000.关于北太平洋气候体制变化的性质和影响的国际气候研究中心专题讨论会的介绍。raybet雷竞技最新海洋科学进展47(2-4):99-102。 哈林顿,j.a., R.S. Cerveny和R.C. Balling, Jr. 1992。南方涛动对北美的影响西南季风.自然地理13:318-330。 黑斯廷斯,j.r.和R.M.特纳,1965年。不断变化的里程。干旱半干旱区下一英里植被随时间变化的生态学研究。亚利桑那大学出版社,图森。 海登,1998年。景观尺度上生态系统对气候的反馈。raybet雷竞技最新英国皇家学会哲学学报,伦敦B, 353:5-18。 j.t.轩尼诗、R.P.吉本斯、J.M.特隆布尔和M.卡德纳斯1983年。新墨西哥南部牧豆树沙丘和前草原1935 - 1980年植被变化。牧场管理杂志36:370-374。 赫贝尔,c.h., F.N.阿瑞斯和R.A.赖特,1972。干旱对草原的影响。生态53:1084 - 1094。 希金斯,r.w., Shi W. 2000。导致美国西南部夏季风年际变化的主要因素。气候变化学报,13(4raybet雷竞技最新):759-776。 霍尔布鲁克,n.m., M.A. Zwieniecki, 1999。栓塞修复和木质部张力:我们需要奇迹吗?植物生理学120:7-10。 r.d.霍尔特和T.H.凯特,2000年。范围限制的其他原因:元种群视角。生态学通讯3:41-47。 汉弗莱,1958年出版。沙漠草原。植被变化的历史及其原因分析。植物学评论24:193-251。 汉弗莱,1987年出版。90年535英里:墨西哥边境的植被变化。新墨西哥大学出版社,阿尔伯克基。 卡亚,E.和J.A.德拉库普,1993。与厄尔尼诺有关的美国水流模式Niño/南方涛动。水资源研究29:2491-2503。 J. E.凯默,P.A.马凯特,J. x .维拉斯科-埃尔南德斯和S.A.莱文,2000。动态景观中的灭绝阈值与元种群持久性。美国博物学家156:478-494。 拉提夫,M.和T.P.巴奈特,1996。北太平洋和北美的raybet雷竞技最新年代际气候变化:动态和可预测性。气候杂志:2407-2raybet雷竞技最新423。 林顿,m.j., J.S.斯佩里,D.G.威廉姆斯,1998。刺柏和毛松水分输送的限制:对耐旱性和蒸腾调节的影响。功能生态学12:906-911。 曼盖,1994年。沙漠化:自然背景与人类管理不善。Springer-Verlag出版社,纽约,314页。 新泽西州曼图亚和S.R.黑尔,2002年。太平洋年代际振荡。海洋学报58:35-44。 Martínez-Vilalta, J.和W.T. Pockman, 2002。奇瓦瓦沙漠三叶大孔草对冻害木质部空化的脆弱性。美国植物学杂志89:1916-1924。 麦凯布,g.j.和M.D.迪丁格,1999。ENSO强度的年代际变化与美国西部降水的远程联系。国际气候学杂志19:1069-1079。 g.r.麦克弗森,H.A.赖特和D.B.韦斯特,1988。半干旱德州草原上灌木入侵的模式。美国米德兰自然学家120:391-397。 米诺贝,1997。北太平洋和北美50-70年的气候振荡。地球物理研究快报24:683-686。 米诺贝,1999。北太平洋双年和十五年气候振荡的共振:在气候体制转变中的作用。raybet雷竞技最新地球物理研究快报26:855858。 米切尔,V.L., 1976。美国西部气候的分区raybet雷竞技最新应用气象学杂志15:920-927。 莫勒斯,小m.c.和C.N.达姆,1990。El Niño和La Niña:对河流生态的全球影响。北美底栖学会杂志9:68-76。 小莫尔斯,C.N.达姆和M.T.克罗克,1992。半干旱地区的气候变化和河流。R.D. Robarts和M.L. Bothwell,编辑,半干旱地区的水生生态系统:对资源管理的影响。加拿大环境部,萨斯卡通,加拿大。 尼尔森,R.P. 1986。高分辨率气候分析与西南生物地理。科学232:27-34。 帕尔默1965年气象干旱.研究论文第45号,美国商务部气象局。华盛顿特区。 帕克斯,j.a., J.S.迪恩和J.L.贝当古,出版,17世纪70年代新墨西哥中南部的年轮、干旱和普韦布洛遗弃。D.E. Doyel和J.S. Dean,编辑,古代西南地区的环境变化和人类适应。犹他州盐湖城犹他大学出版社。 彭,c.s., N.布拉斯,2000。多油藏系统流入量的实际估算。水资源规划与管理学报,asce 126:331-334。 w.t.波克曼和J.S.斯佩里,1997。冻育诱导的木质部空化与三叶大穗的北缘。环境科学109:19-27。 w.t.波克曼和J.S.斯佩里,2000年。索诺兰沙漠植被的空化易损性和分布。美国植物学杂志87:1287-1299。 雷德蒙,k.t.和R.W.科赫,1991。美国西部的地表气raybet雷竞技最新候和水流变率及其与大尺度环流指数的关系。水资源研究27:2381-2399。 雷根斯伯格,1996年。1953-1961年新墨西哥州干旱、牧场和政治的一般动态。新墨西哥历史评论,1月:25-49。 施莱辛格,w.h., J.F.雷诺兹,G.L.坎宁安,L.F.休内克,W.M.贾雷尔,R.A.弗吉尼亚和W.G.惠特福德,1990。全球荒漠化生物反馈,科学247:1043-1048。 施耐德,N.和A.J.米勒,2001。预测北太平洋西部气候。raybet雷竞技最新气候杂志:3997-4raybet雷竞技最新002。 斯佩里,j.s.和M.T.泰利,1988。的机制水的压力诱发木质部栓塞。植物生理学88:581-587。 斯佩里,j.s., K.L.尼科尔斯,J.E.M.沙利文和S.E.伊斯特拉克。1994。犹他州北部和阿拉斯加内陆的环孔、扩散孔和针叶树的木质部栓塞。生态75:1736 - 1752。 t.w.斯威特南和J.L.贝当古,1990。美国西南部的火-南方涛动关系。科学249:1017 - 1020。 t.w.斯威特南和J.L.贝当古,1998。美国西南部年代际气候变化的中尺度扰动和生态响应。气候杂志11:3128raybet雷竞技最新3147。 托马斯,1963年逝世。西南地区干旱影响的总总结:1942-1956年西南地区干旱。美国地质调查专业论文372-H。 m.t.泰里和J.S.斯佩里,1989年。木质部易空化和栓塞。植物生理与分子生物学回顾40:19-38。 瓦伦丁和J.J.诺里斯,1964年。新墨西哥南部选定木酚灌木遗址土壤的比较研究。牧场管理杂志17:23-32。 维拉巴,R.D.达里戈,E.R.库克,G. c .雅各比和G.怀尔斯,2001。沿美洲热带外西海岸的十年尺度raybet雷竞技最新气候变化:来自树木年轮记录的证据。编辑Vera Markgraf,《半球间气候联系》第155-172页。raybet雷竞技最新学术出版社,圣地亚哥。 威廉姆斯,j.e., S.D.戴维斯和K.波特伍德,1997。火后束状腺瘤幼苗的木质部栓塞和复芽。澳大利亚植物学杂志45:291300。 j.a. Yeakley, R.A. Moen, D.D. Breshears和M.K. Nungesser, 1994。北方的回应 美国生态系统模型与温度和降水的多年周期性景观生态学9:249-260。约克,j.c.和W.A.迪克-佩迪。1969.新墨西哥南部植被在过去一百年间的变化。W.G. McGinnies和B.J. Goldman编辑,《干旱区透视》第155-166页。亚利桑那大学出版社,图森。 继续阅读:道格拉斯·G·古丁·莫里斯·J·麦克休 这篇文章有用吗?
 values and the precipitation record reconstructed from the Arroyo de Milagro tree-ring data.")
 contiguous with Chihuahuan Desert vegetation on the south-facing slope (right).")
干旱的影响
 and grasses (Sporobolus spp.).")
 and Larrea tridentata (filled dots) with respect to cumulative precipitation over the 394-year surrogate precipitation history from Arroyo de Milagro, Sevilleta LTER.")
冷冻效果

结论
总结
参考文献
推荐项目



读者的问题
-
沃尔夫冈3个月前
- 回复
-
肯塔基州RYYN3个月前
- 回复