食叶虫
简单的力量
1。介绍
岛屿是深刻有趣的生态系统来研究进化模式和流程。隔离和简单提供自然“Ecotrons”中寻求理解的角色功能的生物多样性的生态系统。孤立的生物群通常relictual,发育不全的和不谐和(集集大1994)和生物多样性显著低于相当于大陆的栖息地(惠塔克1998)。麦克阿瑟和威尔逊(1967)岛屿生物地理学理论也表明,连续的移民和岛屿上物种灭绝导致的动态平衡常数物种周转率。毫不奇怪它已成为公理代表岛屿生态系统简单,脆弱,尤其是容易流行的生物多样性的丧失后入侵大陆生物群(麦克阿瑟和威尔逊1967年集集大1994年,惠塔克1998年Primack 2002)。虽然经验不是很支持(威尔逊2001),平衡理论存在的吸引人的逻辑,甚至被应用于昆虫:植物相互作用(Feeny简森1968,奥普莱1974年,1976年)。然而,壮观的散度岛生物群以来一直认可华莱士(1858)和《达尔文》(1859)。集体、岛屿持有更大的生物多样性比大陆和岛屿植物的不成比例的地方的特性,和副无脊椎动物区系密切相关,表明进化时间尺度的稳定性和可预测性。
共同进化和可预见性的基础是我们目前所了解的昆虫:植物交互康奈尔和霍金斯(2003)。的解释次要的植物化合物,和昆虫微型Dethier(1954)和Erlich设计和乌鸦(1964)主张一个动态的进化,在相互适应,选择植物产生新颖的化合物作为防御食草动物,食草动物开发defense-catabolizing机制在回应。芬尼(1975、1976)的主机可见性的概念试图进行分类植物次生化合物和解释他们的植物区系分布关于社区
t·d·佩恩(主编),入侵森林昆虫介绍了森林树木,1-13和改变生态系统。©2006施普林格。
植物的状态。Southwood(1961)已经演示了一个地理范围之间的正相关关系,或寄主植物出现的可预测性,相关的食草动物的多样性为英国的树木。芬尼认为植物更明显,通过丰富或长寿,必须适应这个大食草动物的多样性。他推断,伍迪高潮的物种,这常常发生在几乎单一的社区,应该采用“定量防御”,如单宁和叶子的韧性,对各种食草动物提供保护。这些非特异性威慑提供转让防御,减少他们的消费比例的树叶可消化性。相反,他认为,不那么明显的植物可能逃脱食草性在空间和时间,或被定性为“独特的毒素浓度相对较低,这可能只被食草动物专家反驳道。共同进化的融合和可见性假说被更进一步的莱文(1975),预测植物抵抗无脊椎食草动物将最好的在中心,寻求最大的可见性的面积,植物的地理范围。然而,地理变异在植物抵御食草动物还没有被很好地解决经验。
简森(1975),莱文(1976年),后来穆迪(1978)认为,生物碱的纬向分布的负相关,其他次要的植物化合物,是放牧压力的回应。舞毒蛾(Lymantria dispar)生物米勒和汉森(1989)产生的证据表明热带树木更好保护比温带和文学评论(Coley &巴龙1996年,戴尔& Coley 2002)支持这一结论。Carlquist(1974)描述了夏威夷群岛是非常可怜的有毒植物和食草动物的建议放松压力群岛导致的损失防御岛植物中的化学物质。加拉帕戈斯群岛的亏损cyanogenesis植物也被解释的食草动物减少压力(Adersen et al ., 1988),和一般损失岛植物防御脊椎动物食草动物的报道鲍恩和van Vuren (1997)。
生物地理学的集体智慧,共同进化,可见性和生物多样性假说预测,简单的,温和的,孤立的植物应该表现出一种固有的脆弱性入侵大陆,r-selected多面手。为什么那么温和,岛,新西兰森林植物明显抵抗入侵,杂食性的,大陆Macrolepidoptera吗?
2。新西兰食叶害虫的经验
有些人会认为新西兰是一个多岛屿。然而,65年的孤立和冰川作用和海洋过犯有时,这限制了特有的纤细的聚集地,当下流行生物群有限状态被认为是发育不全的连岛标准(2000年劳顿,Rosenzweig珀耳斯。通讯)。沃森(2002)认为性格之间的差异通过地理分隔一个生物群,一个由长距离传播,会失去,在这样一个时期。然而,海洋的争论可能的后果很小,为论文提供了这里似乎同样适用于大陆群岛,或任何植物种群,是孤立的,即使同属的包围。
2.1。与生物入侵鳞翅目
Lymantriids没有出现在新西兰特有的无脊椎动物和一连串的机构和拦截Lymantriids (Orgyia thyellina, Teia anartoides, Lymantria dispar)和入侵arctiid Hyphantria cunea,在过去的十年里应该导致容易入籍的森林食叶虫。根除相当大的项目是为了防止这种(霍斯金表示:et al ., 2003)和支持的巧合生物入侵者评估风险对归化和特有植物。所有的生物分析方法之后Matsuki et al。(2001)。在没有选择随机区组试验参数的幼虫死亡率和新生儿的发展记录直到死亡或蛹化。
2.1.1。大陆和岛屿植物之间的区别
凯生物(2002、2003、2004;凯et al ., 2000;Matsuki et al ., 2001;霍斯金表示:et al ., 2003)展示了一个令人惊讶的程度的抵抗这些流行菌群内的食叶害虫。此外,南半球的新西兰代表高潮森林属、假山毛榉(山毛榉目:Nothofagaceae),在很大程度上令人不快的这些奇异的食叶害虫。
假山毛榉物种是长寿的,高潮的优势种流行南半球温带森林,通常发生在“可预测”单一的森林。最伟大的物种多样性和地理范围属发生在南美洲。在一个明显的悖论,生物分析显示,假山毛榉的南美大陆的物种,它受到的影响更大的多样性比新西兰代表无脊椎动物(McQuillan 1993;奥格登et al ., 1996;维布伦et al ., 1996),通常是更容易比新西兰。一些南美假山毛榉更美味的分析比北半球的主要寄主植物鳞翅目!
2.1.2。纬度和假山毛榉国防
如果植物防御纬度呈负相关,在人们的预料之中较低的纬度假山毛榉辩护比更高的纬度物种。这并不是证实假山毛榉的南美洲。最北部的物种n obliqua (33°-41°S)是所有生物中最美味的明显。然而,n . alessandri (35-36°S)一小(350公顷)在同一纬度relictual人口是最美味的物种(Russell et al ., 2000;凯2002;Matsuki et al ., 2001)。此外,n . obliqua最大数量的无脊椎动物和n . alessandri最相关(McQuillan 1993)。澳大利亚n . moorei (28-32°S)是最北部的温带假山毛榉,但也是非常有限的分布。这是最美味的澳洲物种凯(2002)。假山毛榉alessandri和n . moorei可能被认为是大陆的岛屿假山毛榉,这样看来,有限的地理范围的一个物种一样内陆重要的植物防御岛屿上。 It would also appear that area is more important than latitude in determining plant defense.
纬度的复利效应可能在生物分析的产地n . truncata新西兰最美味的山毛榉的物种。假山毛榉truncata是最常见的新西兰山毛榉和有一个小(ca 40公顷)南部人口在哈斯特(44°S),孤立的南部约260公里的巨大人口的物种(Mark &李1985)。与t . anartoides生物测定和两个产地的n . truncata最南端,哈斯特出处证明完全耐药(幼虫死亡率100%)而幼虫在一些复制的Mangorewa (38°S)出处,接近中心的地理范围的n . truncata幸存生产雄蛹(图1)。
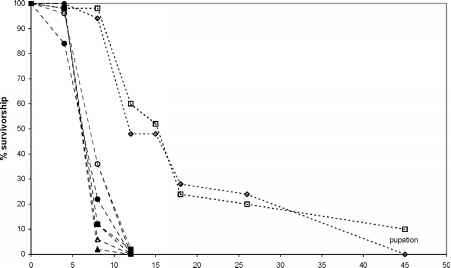
25天
图1所示。的生存Teia anartoides美联储在哈斯特(固体)和Mangorewa(空心)产地ofNothofagus truncata。
25天
图1所示。的生存Teia anartoides美联储在哈斯特(固体)和Mangorewa(空心)产地ofNothofagus truncata。
2.1.3。相应的测试
年辐射动物是主要人工林物种在新西兰。松果体是几乎完全局限于北半球的地理范围
P。放射虫纲是限于两三个离散的大陆和岛屿人口,沿着加州海岸。
自然种群的生物测定,树叶p .放射虫纲,生长在一个共同的花园在新西兰,新西兰土著尺蠖的美联储,Pseudocoremia suavis。幼虫生长速度是贫穷的两个岛上产地p .放射虫纲和增长之间的正相关,个人对加州产地的地理范围(图2)。岛上种源最美味,而且纬度最低的产地。起源和最小的地理范围(Cedros岛)是最美味的。有趣的是新西兰长白猪,第一代商业选择的增长,是最美味的。
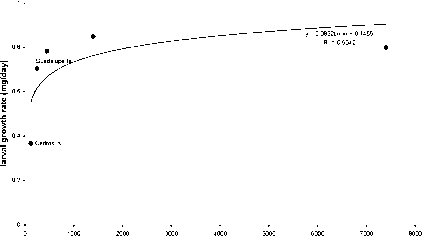
地理范围(公顷)
图2。增长率o / Pseudocoremia suavis美联储在松果体辐射动物从地理上不同的自然种群。
地理范围(公顷)
图2。增长率o / Pseudocoremia suavis美联储在松果体辐射动物从地理上不同的自然种群。
3所示。新西兰的解释经验
岛上资源分配(IRA)假说(凯& Wratten 2004)被开发作为一种解释的表观电阻新西兰植物新食叶虫。爱尔兰共和军的基础假设是一个重新定义的基本生态原则的物种:面积的关系。与区域物种多样性增加,但更明显的关系在更高营养级(霍尔特1996)。栖息地的分裂研究(Kreuss & Tscharntke 1994 schoen 1989年,2000年劳顿,Komonen et al ., 2000)和营养级模式在空间上离散的生态系统(Schoenly et al ., 1991;霍尔特1996)表明,无脊椎动物社区的营养成分不均匀变化区域。相反,在栖息地的分裂,较高的营养级过早丢失或不成比例,或无法维持在小的栖息地。在重新定义物种:区域概念,爱尔兰共和军假说认为,岛屿,或者类似的地理限制生态系统,支持生物多样性较低,具有更少的营养水平,因此有一个较小的自上而下的监管食草动物的天敌。假说认为如果受到食草动物植物健身,然后,在自上而下的食草动物监管薄弱由于缺乏营养的复杂性,植物无法逃避选择在时间或空间上分配资源以自下而上的防御。如果营养多样性促使资源配置的植物防御,降低生物多样性典型的高纬度地区可能复合的物种:区域的影响。
的分手Gondwanan大陆是一个分裂的假山毛榉macroecological规模的生态系统,在进化的时间。温带生态系统存在的残余在澳大利亚、新西兰和南美洲。新西兰大陆的隔离和洪水导致一个发育不全的冷云生物群缺乏营养的复杂性。在大陆类似的气候事件被边缘化的一些物种,这样他们就像大陆的岛屿。来证明这些生物地理限制物种能很好地防御食叶虫。
4所示。讨论
4.1。爱尔兰共和军的假设
人们普遍认为,由于食草动物选择压力,植物进化出了大量的物理、化学、物候和共生战略提供一个自底向上的限制植物的可食叶害虫(爱德华兹& Wratten 1980)。这些策略可能会辅以天敌的行为,影响一个自上而下的监管食草动物(海尔斯顿et al ., 1960)。这一监管多元化现在被认为是存在于大多数系统(猎人&价格1992),但是经验很难证明相对每个琼斯(Walker & 2001)的角色。
与简化的研究趋势似乎有一些实证试图把植物防御策略到一个社区或生态环境。大多数文献昆虫和植物之间的交互忽视随之而来的交互与其他营养级。任务经验几乎是不可能的。即使这样的造型相互作用被认为是令人生畏的复杂性(Holt & Loreau 2001)而不切实际的如果简化(城邦1991)。“绿色世界”假说(海尔斯顿et al ., 1960)认为植物防御的必要性是最小的自上而下的监管影响天敌的食草动物。相反,并不是所有的植物都是食草动物。食草动物的植物防御可能会提供一个自底向上的限制人口1978(白色)。似乎逻辑,如果昆虫取食植物影响健康,那么,当一个健壮的自上而下的监管食草动物种群的天敌是在进化过程中减弱或消失,植物无法逃避在时间或空间上应选择将资源分配给一个“自下而上”的防御。生物多样性在生态系统稳定性的作用是一个热议的问题(Kinzig et al ., 2001),但爱尔兰共和军假说认为,生态系统的稳定性可以通过和解的自顶向下和自底向上流程主要由生物多样性通过介导栖息地的面积。这似乎是反直觉的,只是因为历史悠久,但不支持,假设生物地理学和可见性。
芬尼的防御植物化合物分类变得模糊。芬尼(1975)自己认为对定量和定性策略比较适口性高潮植物物种和英国的树木还没有令人信服地证明(爱德华兹et al ., 1986)。最近定性防御系统已经被证明在存在剂量依赖的相关性定量方式(Lindroth &卷边1990)和植物单宁的作用一直是许多研究的主题,显示一个伟大的和多样的反应。不用说,生物不同,单宁是一种极其变量组化合物和可见性假说坚持(Coley 1983;Forkner et al ., 2004)。爱尔兰共和军的假设会得出这样的结论:可见性假说的极性应逆转。分散植物受食草动物的过度的一篇网络复杂的营养。地理位置限制植物需要固有防御食草动物,需要增加作为栖息地面积减少即“unapparency”假说的植物防御可能更适合。
推论的可见性(芬尼1976)和植物共同进化假说认为孤立与non-evolving防御化合物很容易克服通过食草动物选择压力,和相关的植物应该有类似的二次化学反应。二次描述生态学的植物化学物质还没有被很好地解决了然而,玻姆的岛(1998)有限公司审核二级植物化学表明,岛屿植物化学概要文件有点矛盾。他们可能是相似的,简单的或丰富,相比,相当于大陆物种的概要文件。新西兰植物(80%)高程度的特有现象(Allan 1982)可能提供一个新颖,将阻碍最天主教的喂食器。然而,在众多的证据确凿的情况下新西兰植物丰富的化合物比大陆同行数组。植物系统学并参与化学概要文件,但是对于假山毛榉抵抗食叶害虫发生在三个温带亚种和陆地之间,这表明非系统因素影响化学表达式。
莱文(1975)声称,抵抗食草动物应该发现植物的地理范围的中心,因为更大的食草动物的多样性,忽略了天敌的影响将伴随食草动物。爱尔兰共和军的假说,包括社区营养的复杂性,表明电阻应寻求在孤立的种群或人群的边缘。植物范围的扩张和收缩由于气候或竞争将产生“边缘人群”美国斯特恩和罗氏公司(1974),充当“大陆”的岛屿。raybet雷竞技最新这些孤立的人群选择极端的气候和竞争的物种。raybet雷竞技最新似乎,已经证明为假山毛榉和松果体放射虫纲,这些边际人群也选择防御食草动物。类似的阻力在人群的边缘已经在白松象鼻虫之间的交互和云杉在加拿大(Alfaro et al ., 1999)。
生物活性化合物在植物叶子可能会透过食物链,除非异化的植物叶片衰老。有趣的是,瓦尔德et al .(1997)发现,次生植物化合物的积累与反垃圾增加面积的岛屿高纬度斯堪的纳维亚湖泊。区域生物多样性的争议是最好的预测(1995年Rosenzweig, 1997)。如果生物多样性在植物防御的部署中介,高纬度和高海拔种群的物种也可能揭示抗植物。
4.2。纬度:温带和热带森林如此不同?
纬向梯度生物多样性是一些最古老的观察出版生态学(华莱士1878年)。假山毛榉森林的证据显示明显差异的普遍提倡的思想纬度和食草动物的部署防御。纬度较低的物种:obliqua: alessandri和n . moorei区域,或植物的地理范围的程度,似乎比纬度有更大的影响。n . truncata和p .放射虫纲纬度的影响可能加剧了低面积:生物多样性的影响。
热带森林明显不同于北方或温带地区,它们包含大量的植物物种,而不是更多的单特异性高纬度地区的森林。是在这样一个多元化的热带环境中个体植物“岛屿”的,理智简森(1968)和奥普莱(1974),和为自己辩护,尽管总体天敌的多样性?热带森林的空间复杂性可能使监管食草动物的天敌更加困难。食草动物的自下而上的限制,似乎在海洋和大陆经营岛屿外推到热带地区个别树木吗?
主机专门化通常被解释为共同进化的结果与定性化学防御。提出(1950),Pianka(1966)和麦克阿瑟和威尔逊(1967)推测,食草动物在热带地区的更大的多样性导致了更大程度的主机专业化,使包装更大的物种。专业化已经援引热带无脊椎食草动物的一个特征(代尔& Coley 2002),但它也是一个新西兰的无脊椎动物(Dugdale称1975)。热带地区是次生植物化合物的浓度明显只是因为之前作者没有考虑区域的影响?总是容易分析的常见代表温带属或家庭,比一个罕见的温带物种和生物测定结果往往是外推到包括同类的(巴博萨& Krischik 1987)。米勒的研究结果和汉森(1989)使用l . dispar生物分析和热带植物没有比温带新西兰例子使用相同的试验。
4.3。增长率和植物防御
一些作者认为国防资源配置是最大的生长缓慢的植物物种在资源贫乏的环境中(简森1974;Coley et al ., 1985;Coley 1988;价格1991;爱马仕&马特森1992)。在新西兰特有的缓慢增长之间的反差森林树木和超级跑车的壮观的增长率是被早期的森林。尽管新西兰几乎完全被森林覆盖的欧洲殖民,木材特有物种的缓慢增长需要切换到异国情调的森林物种在100年创建一个可持续森林产业。外来植物生长在新西兰比在他们的祖国表明新西兰特有的不受到资源的限制。是他们的增长速度缓慢的资源分配的结果防御,而不是需要一个防守,因为他们长得这么缓慢?几乎没有的比较为假山毛榉增长数据。 However, New Zealand species failed to thrive in the experimental plantings of the genus in UK, whereas the South American species N. obliqua (the most palatable species) has potential for plantation forestry there (Tuley 1980). It is also interesting to note that New Zealand commercial selections of Pinus radiata grow more rapidly than the original Californian populations, but are considerably more palatable to invertebrate defoliators. The artificial selection for growth appears to have been at the expense of resource allocation to defense.
5。结论
复杂的食物网动力学限制的经验综合生态形状组件社区内物种进化的力量。事情的真相是,很少有昆虫:植物的研究包括tritrophic交互,除了植物防御化学的影响健康的天敌。过多的假设已经开发了特定的相互作用,但没有最重要的假设,实证测试,适合所有。爱尔兰共和军的假设是可测试的,提供了一个“红移”证明生物多样性决定了生态系统功能,没有恢复到人为ecotrons似是而非地简化模型或昂贵。有悖于传统智慧,岛上森林抵抗入侵大陆食叶害虫出现。明显不同的部署防御无脊椎动物食草动物已经证明与同属的不同地区的同种的植物在本质上相同的栖息地。岛森林似乎有一个固有的力量在岛社区的简单。区域生物多样性和营养的主要决定因素的复杂性。爱尔兰共和军的解释之间的交互高潮森林和食叶害虫驳斥了建立假设。相反,它认为任何森林食叶害虫的互动是由自顶向下和自底向上的相对优势力量决定系统,这是由栖息地面积,和栖息地面积可能是海洋或大陆,和温带或热带物种。
新西兰森林研究所
私人包3020
罗托鲁瓦
新西兰
Ph值+ 64 7 343 5500
电子邮件(电子邮件保护)
6。引用
Adersen,。Adersen, H。&布莱墨,l (1988)。生氰成分在植物从加拉帕戈斯群岛
岛屿。生物化学分类学和生态学,16岁,65 - 77。居多,e·汤姆林,J.A. Borden & k·刘易斯。(1999)。交互的白松象鼻虫和它的主人:共同进化的观点。31 - 39页。f . Lieutier, j•马特森,核磁共振瓦格纳(Eds)、生理学和遗传学的tree-phytophage交互。Gujan学报IUFRO会议,法国。8月31-Sept。5,1997。INRA、巴黎。374页。艾伦,h . h (1982)。 Flora of New Zealand. Vol.1. Wellington : Hasselberg, Government Printer, Barbosa, P., & Krischik, V. A. (1987). Influence of alkaloids on feeding preference of eastern deciduous forest trees by the Gypsy moth, Lymantria dispar. American Naturalist, 130, 53-69. Bohm, B. A. (1998). General evolutionary patterns and processes on oceanic islands. In T.F. Stuessy &
m .小野(Eds),岛屿植物的进化和物种形成。剑桥大学出版社。鲍恩e . & van Vuren d (1997)。岛特有植物缺乏抵御食草动物。
保护生物学,1249 - 1254。Carlquist,美国(1974年)。岛生物学。纽约:哥伦比亚大学出版社。
绿青鳕,P.D. (1983)。食草性和防御性低地热带森林树种的特点。
生态专著,53岁,209 - 233。绿青鳕,p D。,科比,j . p . &查宾f s (1985)。资源可用性和植物antiherbivore防御。
科学,230,895 - 899。
绿青鳕,p . d . (1988)。影响植物的生长速率和叶一生antiherbivore数量和类型的防御。环境科学,74,531 - 536。绿青鳕,j·p·d·&巴龙a (1996)。食草性和在热带森林植物防御系统。年度回顾的
生态学和系统学,27岁,305 - 335。康奈尔,b . a . h . v . &霍金斯(2003)。食草动物对植物次生化合物:植物化学的共同进化理论的一个测试。美国博物学家,161;508 - 522。达尔文,c (1859)。在《物种起源》通过自然选择或保存青睐的斗争中生活。伦敦:约翰·默里Dethier诉g (1954)。喂养喜好食植物的昆虫的进化。提出进化,8日,t (1950)。 Evolution in the tropics. American Scientist, 38, 209-221
Dugdale称,j . s . (1975)。的昆虫与植物的关系。在g . Kuschel。生物地理学和生态学
新西兰。Monographiae Biologicae 27日海牙:垃圾,戴尔,洛杉矶& Coley p d (2002)。Tritrophic交互在热带和温带的社区。在T。
Tscharntke & b·a·霍金斯(Eds) Multitrophic水平交互。剑桥大学出版社爱德华兹,p . j . & Wratten s d (1980)。生态insect-plant交互。121年生物学研究。伦敦:爱德华·阿诺德
爱德华兹,p . J。Wratten, s d &格林伍德,美国(1986年)。英国树的适口性昆虫:
本构和诱导防御。环境科学,69,316 - 319。Erlich设计,p . r . &乌鸦,p h (1964)。蝴蝶和植物:共同进化的研究。586608年进化,18日。
芬尼,p (1975)。生化共同进化的植物和昆虫之间的食草动物。L.E.吉尔伯特&
由P.H.乌鸦(Eds),共同进化的动物和植物。奥斯汀:德克萨斯大学出版社。芬尼,p (1976)。植物可见性和化学防御。最近的植物化学的进步,第1 - 40 10日。Forkner r E。侯爵,r . j . &莉儿,j . t . (2004)。芬尼的再现:浓缩单宁作为leaf-chewing anti-herbivore防御食草动物Quercus的社区。生态昆虫学,29岁,174 - 187
Futuyma, d . j . &米特(1997)。Insect-plant交互:组件社区的进化。:Silvertown, j .;弗朗哥,m;哈珀,j·l . (Eds)植物生活史(页253 - 264)。英国剑桥大学出版社。
海斯:G。f·e·史密斯& Slobodkin l . b . (1960)。社区结构、人口控制和竞争。美国博物学家,94年,421 - 425。
爱马仕。d . a &马特森,w . j . (1992)。植物生长或保护的困境。生物学评论季刊》67年,283 - 335。
霍尔特,r . d . (1996)。在太空食物网:一个岛屿生物地理的视角。在G.A.城邦和快速出拳Winemiller。(Eds)、食物网。集成模式和动力学(页312 - 323)。纽约:查普曼和大厅,
霍尔特,r D。劳顿,j . H。城邦,g。&马丁内斯,n . d . 1999。营养等级和物种-区域关系。生态学,80,1495 - 1504。
霍尔特,r·d·& Loreau M。,(2001)。生物多样性和生态系统功能:营养的角色交互和系统开放的重要性。在a . p . Kinzig s w·帕卡拉& d Tilman (Eds),生物多样性的功能的影响:实证研究的进展和理论扩展(页246262)。普林斯顿:普林斯顿大学出版社,
霍斯金表示:,他清水,J。Handiside, J。凯,M。雷j . Simmons & n (2003)。毒蛾根除——一个来自新西兰的成功故事。,现年49岁的害虫管理的国际期刊1724年。
猎人,m D。&价格p w (1992)。玩降落伞和梯子:异质性和相对角色自顶向下和自底向上的力量在自然的社区。生态学,73,724 - 732。
简森,d·h·(1968)。宿主植物岛屿在进化和当代的时间。美国博物学家,102年,592 - 595。
简森,d·h·(1974)。热带黑水河流,动物和桅杆果期龙脑香料。
时,69 - 103。
简森,d·h·(1975)。植物在热带地区的生态。研究生物学。58岁的伦敦:爱德华·阿诺德。
凯,m . k . (2002)。调查的适用性新西兰森林植物Lymantriid物种的繁衍和发展。Unpubl。MAFResearch报告,42页。
凯,抗议(2003)。Macroecology和入侵无脊椎动物协会的预测。Goldson S &乳儿,m . (Eds)保卫绿色绿洲:新西兰生物和科学。《植物保护学会学报新西兰。生物安全研讨会,罗托鲁瓦,2003年
凯,m . k . (2004)。的风险评估对新西兰植物造成日本舞毒蛾的菌株和web-worm下降。Unpubl。我们研究报告,29页。
凯,M。Matsuki, M。金丝雀,J。& Scott, J.K. (2000). A risk assessment of the Asian gypsy moth to key elements of the New Zealand flora. Unpbl. MAF Research Report 82pp.
凯,m . k . & Wratten s d (2004)。生态系统功能和树抵抗食叶害虫的预测。。在n Kamata(主编),诉讼21世纪COEProgram金泽大学的国际研讨会。11. 15.11.2003 Kanzawa大学,日本媒体
Kinzig, a P。d . s . w . &,帕卡拉·蒂尔曼(Eds) (2001)。生物多样性的功能结果。人口生物学专著33岁的普林斯顿:普林斯顿大学出版社。
Komonen,。Penttila, R。林格伦,m & Hanski i (2000)。森林碎片截断食物链基于原始森林托架真菌。Oikos, 90, 119 - 126。
Kreuss, a & Tscharntke t (1994)。栖息地破坏、物种灭绝、生物控制。科学,264,1581 - 1584。
劳顿,j . h (2000)。社区生态变化的世界。卓越在生态11中,德国:生态研究所。
莱文,d . a (1975)。在植物害虫的压力和复合系统。美国博物学家,109年、437451年
莱文d . a (1976)。Alkaloid-bearing植物:ecogeographical视角。美国博物学家,110年,261 - 84
Lindroth, R.L.,& J.D.C.卷边。(1990)。反应的舞毒蛾(鳞翅目:Lymantriidae) tremulacin,白杨树的酚糖苷。环境昆虫学,19岁,842 - 847。
麦克阿瑟,r·h·威尔逊& e·o·(1967)。岛屿生物地理学理论。普林斯顿:普林斯顿大学出版社。
马克,w·g·a·f·& Lee (1985)。生态学的山毛榉(假山毛榉truncata)南部的局外人站在哈斯特生态地区,南韦斯特兰,新西兰。新西兰生态学报,97 - 115
Matsuki, M。凯,M。金丝雀,J。、弗洛伊德、r和斯科特,j·k·(2001)。潜在的意外风险引入亚洲舞毒蛾(Lymantria dispar)大洋洲:气候条件和适用性的本地植物的影响。农业和森林昆虫学,305 - 320。McQuillan, p . b . (1993)。假山毛榉(壳斗科)及其无脊椎动物——概述和初步的合成。生物林奈学会的期刊,49岁,317 - 354。米勒,p . j . c . &汉森大肠(1989)。实验室饲养试验对舞毒蛾幼虫的发展参考植物类群和物质。俄勒冈州立大学农业试验站的公告,674年1 - 63喜怒无常,美国(1978年)。纬度,大陆漂移和alkaloid-bearing植物的比例组成。
美国博物学家,113年,965 - 72奥格登,J。斯图尔特,g·h·艾伦& r b (1996)。新西兰生态假山毛榉森林。在t·t·维布伦,R.S. Hill & J读(Eds),假山毛榉森林的生态和生物地理学(页2582)。凿天堂:耶鲁大学出版社。奥普莱,p . a (1974)。橡树作为分层开采昆虫进化的岛屿。美国科学,62,67 - 73集集大g (1994)。生物多样性的海洋岛屿上:它的起源和灭绝。美国动物学家,134144年34岁
Pianka e r (1966)。纬向梯度物种多样性:回顾的概念。100年美国博物学家,33-46
城邦,g . a (1991)。复杂的营养相互作用在沙漠:一个实证食物链理论的批判。
138年美国博物学家,价格123 - 155,p . w . (1991)。植物活力假说和食草动物的攻击。Oikos、62、244 - 251 Primack r (2002)。保护生物学的必需品。3日麻萨诸塞州:Sinauer Associates Inc .罗迪斯,测向& Cates R.G. (1976)。植物antiherbivore化学的一般理论。最近
植物化学进展,168 - 213 Rosenzweig, m . l . (1995)。物种多样性在空间和时间。英国剑桥大学出版社。Rosenzweig, m . l . (1997)。物种多样性和纬度:听区域的信号。Oikos, 80, 172 - 176罗素,g B。鲍尔斯,w·S。Keesing, V。尼迈耶,h . M。Sevenet, T。,美国& Wratten Vasanthaverni s d . (2000)。模式生物活性和食草性假山毛榉物种从智利和新西兰。化学生态学学报,26 (1)41-56。 Schoener, T. W. (1989). Food webs from the small to the large. Ecology, 70, 1559-89. Schoenly, K., Beaver, R. A. & Heumier, T. A. (1991). On the trophic relations of insects: a food-web approach. American Naturalist, 137, 597-638. Southwood, T. R. E. (1961). The number of species of insect associated with various trees. Journal of Animal Ecology, 30, 1-8.
斯特恩,k &罗氏l . (1974)。遗传学的森林生态系统。生态学研究6。纽约:SpringerVerlag,
强,d . r . (1992)。都是营养级联湿?分化和donor-control speciose生态系统。生态学,73 (3),747 - 754。强,d R。劳顿j·h·萨斯伍德& r (1984)。昆虫对植物:社区模式和机制。牛津大学:布莱克韦尔科学出版物。Tuley, g (1980)。在英国假山毛榉。 For. Comm. Forest Record 122
维布伦,T . T。Donso C。Kitzberger, a t & Rebertus j . (1996)。智利和阿根廷南部生态假山毛榉森林。r·s·希尔,t·t·维布伦& j .阅读(Eds),假山毛榉森林的生态和生物地理学(页293 - 353)。凿天堂:耶鲁大学出版社。沃克·m·琼斯,t·h . 2001。相对自顶向下和自底向上的角色在陆地的三倍
营养plant-insect-natural敌人系统。Oikos, 93, 177 - 187。华莱士a r (1855)。法律监管的新物种的引入。一年一度的自然历史》杂志,184 - 196。华莱士,a . r . (1878)。热带自然和其他论文。纽约:麦克米伦。
瓦尔德,d . A。Zackrisson, O。公司,G。&石片,c (1997)。岛地区生态系统特性的影响。277年科学,1296 - 1299。沃森·d·m . (2002)。一个概念性的框架研究物种组成的群岛,碎片和其他的栖息地。29岁的生物地理学杂志823 - 34。
白色,t . c . r . (1978)。一个相对的重要性食物短缺在动物生态学。环境科学,33岁,71 - 86
e·o·威尔逊(2001)。前言版本2。岛屿生物地理学理论。麦克阿瑟,r·h·&
威尔逊,e·o·普林斯顿:普林斯顿大学出版社。惠塔克,r . j . (1998)。岛屿生物地理学。牛津大学。
这个页面故意空白
继续阅读:本地昆虫殖民介绍树Speciespatterns和潜在风险
这篇文章有用吗?