X
鞭毛虫
的p jahnii鞭毛虫是圆的,直径3.5 - -5.0 |米时观察到的生活,和有两个到四个金壁叶绿体。熊不平等的两个长鞭毛(8.5 | -12 m和5.56.5 |米),一个non-coiling haptonema 3.0 | -4.5米的长度,和两个规模类型(Zingone et al . 1999年)。丝没有观察到。
p . cordata的形态类型
棕囊藻属cordata但是此属只有被描述为一个鞭毛细胞(Zingone et al . 1999年)。活细胞是3 - 3.5 |米长,3 - 4 |米宽。他们有两个不相等地长鞭毛的5.5 - -7.5 m和4.5 | 6 |米的长度,和non-coiling haptonema 2.2 | -2.5米长。他们拥有两个类型,规模与微弱的辐射山脊。这些细胞产生细丝形成五角的数字。
p . scrobiculata的形态类型
棕囊藻属scrobiculata,但是此属已观察到偶尔在样品和从未在文化领域,只是报道鞭毛虫(Moestrup 1979;Hallegraeff 1983;Estep等等。1984;Chretiennot-Dinet未公开的数据)。的
-
")
- 图3的示意图表示haploid-diploid生命周期(从瓦莱罗能源et al . 1992年)
")
细胞、8 |米长度与四氧化锇固定时,熊两个平等的鞭毛和non-coiling haptonema。覆盖的periplast两种类型的尺度,并产生细丝超过50 |米形成图而不是five-ray nine-ray明星中观察到其他棕囊藻属物种,但是此属(Moestrup 1979;Hallegraeff 1983)。巨大差异已报告在鞭毛的长度和haptonema,和身体大小规模(Moestrup 1979;Hallegraeff 1983;Estep等等。1984;Hoepffner和哈斯1990)根据制备过程。
观察到的形态类型的综合
仔细检查文献表明,四种不同的细胞类型中存在Phaeo-cystis物种(表1)。所有六个物种一样,殖民与否,是否发生鳞状鞭毛虫(表1)两种类型。一个恒星形成细丝和所有物种已经报道了p . jahnii除外。,剥夺了细丝,恒星,被发现在p . globosa帕克et al . 1971;Peperzak et al . 2000年)和p . jahnii (Zingone et al . 1999年)。这两种细胞类型是特别重要的,因为他们提供相关分类标准来比较和区分不同属的物种。这些标准包括不同大小、结构和安排的尺度和恒星形成线程(Moestrup 1979;Moestrup和拉森1992;鲍曼et al . 1994;Medlin et al . 1994;Zingone et al . 1999; Jacobsen 2002; Lange et al. 2002). These cells have been shown to be haploid in P. globosa (Vaulot et al. 1994) but diploid in P. pouchetii (Jacobsen 2002).
三个克隆形成物种一样,p . globosa p pouchetii p .南极洲,是三种细胞的发生:鞭打
二倍体鞭毛虫
二倍体鞭毛虫
单倍体鞭毛虫
殖民时期的细胞
年底开花
单倍体鞭毛虫
殖民时期的细胞
年底开花
Inter-bloom:单倍体
布卢姆:二倍性
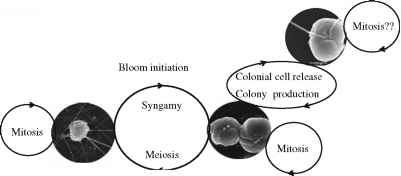
图4 p . globosa haploid-diploid生命周期。单倍体的鞭毛虫的特点是恒星,丝、尺度和大小范围在3.6 - -5.8 m ^当生活。殖民时期的细胞,在尺寸范围5.8 - -10.4 m ^生活,现在两个短顶端附属物,剥夺haptonema,明星、细丝和鳞片。二倍体鞭毛虫,相同大小的范围比殖民细胞,有两个鞭毛,haptonema和缺乏明星,细丝和尺度尺度和细丝,殖民地的细胞,和鞭毛虫缺乏尺度和细丝(表1)。Xagellates尺度和Wlaments殖民之前已经观察到多细胞大量繁殖,当殖民细胞消失,和内部形成大量殖民地的殖民阶段。的存在更大的殖民Xagellates通常局限于发现细胞。在物种并不会形成菌落,p . scrobiculata和p . cordata只有Xagellates尺度和Wlaments观察(表1)。
的棕囊藻属生命周期但是此属
p . globosa haploid-diploid生命周期
不同的存在形态类型两倍性水平与相变有关,和能力的单倍体和二倍体阶段划分mitotically (Kornmann 1955;卢梭et al . 1994;Vaulot et al . 1994年),支持在p . globosa haploid-diploid的生命周期的存在。在这种生命周期,单倍体和二倍体阶段通过性相关流程,减数分裂和两性生殖,都能有丝分裂(图3;
瓦莱罗能源et al . 1992;乌当鸡et al . 2004年)。基于从文化和信息领域的研究提出了在前面的小节中,p的globosa haploid-diploid生命周期已经重建(图4)。这个循环的两个主要突出特点是在殖民地布鲁姆的形成和终止,性是普遍存在两种类型的营养生殖。
棕囊藻属globosa但是此属殖民地开花结果从性过程
单倍体鞭毛虫的发生在两个花朵之间的水柱殖民地二倍体细胞的观察到北海南部(卢梭和Chretiennot-Dinet未发表的数据)提供了证据表明,p . globosa殖民地布鲁姆起始和终止涉及性过程,与二倍体阶段的长度被限制在殖民地花朵。形成一个二倍体non-motile殖民地从单倍体鞭毛虫意味着两性生殖细胞(细胞质和细胞核的融合两个迄今己和随后的受精卵生产)必须发生在殖民地开始绽放。观察两种形态的单倍体鞭毛虫(帕克et al . 1971;Peperzak等等。
2000年)表明,异配生殖发生在p . globosa,但这仍然等待证明。丝的特点单倍体鞭毛虫可以在求偶中发挥作用的。这些结构确实被认为与性(Vaulot et al . 1994年)或在附件中发挥作用(Chretien-not-Dinet 1999)。不能再生dip-loid殖民的单倍体细胞克隆鞭毛虫(帕克et al . 1971;Vaulot等等。1994年)表明,不同的交配类型,即。,兼容的配子形成受精卵,存在于p globosa (Vaulot et al . 1994年)。也不知道是否同宗配合(self-fertile殖民地)或heterothal-lism (self-sterile殖民地)是p . globosa掌权。
相反,减数分裂必须发生在形式hap-loid鞭毛虫从二倍体细胞殖民。这可能发生在单倍体鞭毛虫在殖民地的大规模生产通常报告结束时p . globosa布鲁姆消失前殖民阶段(卢梭et al . 1994;Peperzak et al . 2000年)。然而单倍体鞭毛虫是生产数量限制的殖民地。大多数人开始衰老,随后聚合,被各种异养生物逐步入侵开发复杂的微生物网络(兰斯洛特和卢梭1994;兰斯洛特et al . 2002年)。
在p . globosa营养生殖
的营养生殖二倍体阶段发生在两个不同的途径包括殖民细胞和二倍体鞭毛虫(图4)。一个由殖民细胞的有丝分裂的殖民地,即。,殖民地经济增长(Kornmann 1955;卢梭et al . 1994;Veldhuis et al . 2005年)。这个过程会导致殖民地划分和出芽在昌盛(真实性et al . 1988年),在这个领域(卢梭et al . 1994年),但似乎是次要的。第二个途径包括通过短暂的二倍体鞭毛虫的过渡,从殖民地和释放在一天内能够重新启动殖民阶段(Kornmann 1955;Cariou et al . 1994;卢梭et al . 1994年)。二倍体鞭毛虫因此与殖民细胞共现和殖民阶段,传播途径在实验室常用生产殖民地文化。然而,在自然环境中的意义是非常值得怀疑,可能是由于短寿命缩短的二倍体鞭毛虫(Kornmann 1955;Cariou et al . 1994年)。 The natural occurrence of diploid flagellates in the field is difficult to estimate. It results from disruption of large colonies when the sea is stormy and under turbulent conditions (Kornmann 1955, Peperzak et al. 2000a). Occurrence of these cells, in association with non-motile free-living cells, could however also result from sample manipulation during collection, size fractionation, and incubation. The minor role of the diploid flagellate in the natural environment is also suggested by the observation of massive and synchronous generation of small colonies in the early phase of blooms (Rousseau et al. 1990). A significant vegetative reproduction would instead result in more-continuous colony production. However, this pathway could not be excluded and has been suggested to provide the inoculum for colony blooms (Cadee 1991).
内部因素诱导相变p globosa生命周期
几个因素一直在猜测扮演一个角色之间的转换p globosa形态类型。殖民地的形成从自由细胞磷酸相关损耗(Veldhuis和Admiraal 1987;Cariou et al . 1994年),光强(Kornmann 1955;Peperzak 1993;Peperzak et al . 1998),春天的硅藻产生的化学物质,特别是一些Chae-toceros物种(Weisse et al . 1986;Boalch 1987;卢梭et al . 1994年)和湍流(Schapira 2005;Shapira et al . 2006年)。固体基质细胞附件的要求也被建议从观察小殖民地连着Chaetoceros刚毛在布鲁姆的早期阶段(图1;Boalch 1987; Rousseau etal. 1994; Chretiennot-Dinet et al. 1997).
然而,仔细检查文献表明,大多数研究p globosa集落形成已经完成了实验室培养使用营养生殖途径,即。,培养液二倍体鞭毛虫来自殖民地的中断(表2)。这些研究表明,固体基质,动荡和磷酸因素促进营养代殖民地从字段。(表2)
表2因素参与了从独立生存的细胞转变为殖民阶段p .水珠。 |
阻塞性睡眠呼吸暂停综合症 |
|||
引用 |
因素 |
经验值。 |
独立生存的细胞起源 |
术语和可用 |
继续阅读:对棕囊藻属Globosa但是此属无机碳和有机碳的影响
这篇文章有用吗?