爬行动物的迁移模式
爬行动物迁移的规模急剧变化,从几百米到几千公里。不像两栖动物,爬行动物迁移到一些常见地区繁殖,和最不迁移(齿轮和Zweifel 1998;Pough et al . 1998年)。这是因为爬行动物繁殖不依赖水,因此大多数陆生爬行动物不需要长途跋涉,而一般产卵筑巢地点在其正常活动范围(Pough et al . 1998年)。然而,还有许多其他原因爬行动物的迁移。例如,他们可能在应对季节性迁移的栖息地变化,或跟随他们的猎物。迁移也可能产卵的结果在陆地上,旅行是达到合适,进行地面筑巢地点(Pough et al . 1998年),许多水生爬行动物一样,包括海龟、淡水龟,和一些鳄鱼。其中最广泛研究的迁移是海龟筑巢。
海龟经常承担因迁徙数千公里之间的嵌套和捕食场所(Meylan 1982)(图4)。海龟进行这些长途,或macro-geographic,迁移,因为他们的栖息地通常冷淡地脱离他们的捕食场所(鲍文et al . 1989;林基2001)。不同种类的海龟展览不同的迁徙模式(Luschi et al . 2003年)(图4)。例如,绿海龟(龟鳖目mydas)各殖民地在印度洋和太平洋定期迁移和筑巢海滩觅食区之间1000公里;榄蠵龟龟(摘要本文olviacea)标记在苏里南已发现1900公里;和赤蠵龟(Caretta Caretta)标记在南非曾被观察到在桑给巴尔岛,近3000公里(休斯1974;Pough et al . 2002年)。高游牧棱皮海龟(Dermochelys coriacea)标记在法属圭亚那夺回在各种各样的地方5000公里,包括德州墨西哥、新泽西、新英格兰、新斯科舍省和非洲(Luschi et al . 1996;Pough et al . 2002年)。这些物种的迁徙模式可能是固定与否,取决于他们是否利用可预见或不可预测的资源。然而,海龟迁徙的一个共同特点是,尽管他们遍历的巨大跨越大洋,这些海龟能够返回的精度相同的筑巢海滩每年(Meylan 1982)(图5)。
最深入研究的迁移是绿海龟(龟鳖目mydas)(图4)。这些海龟栖息在热带和亚热带海域,饲料在浅海近岸水域区(卡尔1965;Luschi et al . 1998;无趣,利马et al . 2003年)。每2 - 4年这些海龟穿越大洋,使用快速、定向运动在迁移走廊到达嵌套网站数千公里之外(米勒1998;Luschi et al . 1997;Luschi et al . 2001年)。在加勒比海和大西洋北部,绿海龟利用四个主要筑巢地点位于Tortuguero,鸟类岛,苏利南海岸,阿森松岛。Post-nesting女性迁移数百或数千公里回到觅食区域,虽然有些人可能仍然在筑巢海滩附近一段时间(Luschi et al . 1998;Garduno et al . 2000; Papi et al. 2000; Hays et al. 2001). Females from distantly separated nesting beaches may use the same feeding grounds (LeGall and Hughes 1987).
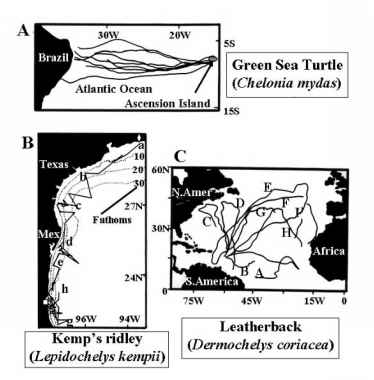
图4所示。代表的海龟迁徙模式。八个提升海龟的路线(龟鳖目mydas)进行越洋穿梭迁移从它们的繁殖地阿森松岛沿着巴西海岸的觅食地。从Luschi修改,海斯和爸爸(2003,图1)。33之间的旅行时间,74天。b .运动satellite-tracked坎普的雷德利(摘要本文kempii)从卡梅隆,路易斯安那州(a,白色钻石)在牧场新筑巢,墨西哥(白色广场)。跟踪时间从1994年8月13日持续(卡梅隆,路易斯安那州——一个)1995年5月16日(h)。时间进程迁移由字母a表示:1994年8月13日;b - 1994年12月10日;c - 1994年12月18日;d - 1995年1月17日;e - 1995年3月8日; f - March 10 1995; g - March 18 1995; h - May 16 1995. Modified from Renaud et al. (1996, Fig. 1). C. Pan-Atlantic foraging movements of nine leatherback turtles (Dermochelys coriacea) tracked after nesting in the Caribbean. Individuals A and B were tracked for 12 months (July 2002 - July
2003)。个人C我跟踪了6 - 8个月(May-July 2003年1月
2004)。修改海斯后,霍顿和迈尔斯(2004,图1)
-
 and Musick and Limpus (1997, Fig. 4.1)")
- 图5所示。概念模型广义海龟的生命周期,Luschi后(2003年,图6.1)和作乐Limpus(1997,图4.1)
 and Musick and Limpus (1997, Fig. 4.1)")
多达50000个龟鳖目mydas上岸来筑巢海滩上Tortuguero,哥斯达黎加每年。成千上万的这些海龟已经标记,和已发现捕食场所远在巴拿马、哥伦比亚、委内瑞拉和尤卡坦半岛(卡尔et al . 1978年)。的生殖电路识别Tortugeura——西部加勒比人口绿海龟(1957年卡尔和Giovannoli)非常类似于其他人群附近也门在印度洋西部(Hirth和卡尔1970),中央南太平洋(Craig et al . 2004),地中海(无et al . 2002年),和中国南部水域程(2000)。
阿森松岛人口的龟鳖目mydas也被广泛研究(图4)。这些海龟迁徙2200公里从巴西海岸的觅食区人口与海龟混合,窝在苏里南和法属圭亚那的海岸,在阿森松岛筑巢海滩(卡尔1962;莫蒂默和卡尔1987)。阿森松岛是仅20公里直径的小岛位于大西洋洋中脊(Pough et al . 2002),且不具备充足的资源来喂养(1974年叛乱)。海龟回到它精确每2 - 3年为了品种和产卵,一段旅程,需要六个星期才能完成(鲍文et al . 1989;Luschi et al . 1998;Pough et al . 2002年)。女性展示地点富达巢,并返回相同的海滩,这通常是他们出生的海滩。这已被证实使用线粒体DNA证据(Meylan et al . 1990年)。
有人建议,海龟的繁殖种群隔离阿森松岛的结果海底扩张(卡尔和科尔曼1974)。这一假说表明,从历史上看,阿森松岛是位于巴西海岸更近,在洋中脊处,海底扩张缓慢向该岛远离巴西,需要阿森松岛龟在越来越大的距离迁移到筑巢海滩(卡尔和科尔曼1974)。然而,检查女性绿海龟的线粒体DNA从三个独立的大西洋聚居地透露,种群之间的遗传差异太小,支持这一假设,而不是显示最近的起源海龟的阿森松岛的人口,可能在过去的几千年中(鲍文et al . 1989年)。这样的问题如何海龟开始筑巢的这个小岛上这仍是个谜。然而,尽管女性从三个聚居地密切相关,互相聚居地是遗传学上截然不同的(鲍文et al . 1989;Avise和鲍文1994)。这表明,女性不仅显示巢网站忠诚,但年复一年,他们回到出生的海滩(鲍文et al . 1989;Avise和鲍文1994)。
东太平洋绿龟的范围(龟鳖目mydas agassizi)仅限于美洲西海岸,从加利福尼亚半岛到秘鲁南部。女性从觅食理由迁移到墨西哥海岸筑巢地点每3 - 4年(阿尔瓦拉多et al . 2003年)。这些海龟是米却肯州的主要筑巢。海龟繁殖后进行回归迁移到加利福尼亚湾的觅食的理由和沿海中美洲和南美洲(Byles et al . 1995年)。
玳瑁海龟(Eretmochelys imbricata)显示一个类似的绿海龟的迁徙模式。这些海龟是广泛分布,通常栖息在珊瑚礁。这个物种最初认为是non-migratory,但遥测法研究表明,玳瑁海龟可能长途迁移(Parmenter 1993;Meylan 1999;Horrocks et al . 2001;Buitrago和Guada 2002)。有证据表明,物种的雄性也高度洄游(Nietschmann 1981)。这个物种的嵌套迁移不是很好理解,但似乎女性回到自己的出生地的海滩巢每2 - 3年(Witzell 1983;低音1999),女性存在于相同的觅食区不迁移到相同的筑巢海滩。玳瑁海龟下蛋后立即迁移回到他们觅食的理由(Horrocks et al . 2001年)。 There is some variation in the length of migration that the females undertake; females nesting on the same beaches exploit different foraging areas that may be either very close to, or a long distance from the nesting beach, and thus migrations may take place across deep oceans or through shallow water (Ellis et al. 2000).
坎皮海龟(摘要本文kempii)表现出不同的迁徙模式的绿海龟和玳瑁海龟(图4 b)。这些海龟有限制范围,在墨西哥湾和大西洋西部和迁移浅水区美国和墨西哥海岸沿着一个定义良好的迁移走廊(Byles和普罗金1994;马尔克斯1994)。成熟女性迁移,总在筑巢海滩在繁殖季节之前,而男性似乎non-migratory,全年保持在筑巢海滩(普罗金2003)。女性每年发生迁移,大多数的坎皮海龟的人口迁移到相同的海滩在牧场新墨西哥(图4 b),虽然有些海龟筑巢在韦拉克鲁斯的几个相邻的海滩,以及在德克萨斯州和其他领域的美国南部(Renaud et al . 1996年)。繁殖后,女性迁移回限量喂养范围(马尔克斯1994;Byles和普罗金1994;Renaud et al . 1996年)。
成年海龟(Caretta Caretta)居住在亚热带和温带水域(Polovina et al . 2004年)在大陆架和河口地区,和绝大多数时间都在海岸和近海水域附近(多德和Byles 2003;无趣,Broderick et al . 2003年)。不像其他海龟物种,雌性海龟不局限于任何一个,固定觅食区,而是一系列之间移动不断觅食区(多德1988;爸爸et al . 1997;普罗金和Spotila 2002)。女性迁移数百到数千公里(Limpus和Limpus 2001;Bentivegna 2002)繁殖地点大约每三年一次,一般在近海水域迁移,只有短暂的海上运动(爸爸et al . 1997;普罗金和Spotila 2002),尽管有些人可能接受跨太平洋沿着特定的迁徙走廊与特定迁移热特性(Resendiz一起做et al . 1988;尼科尔斯et al . 2000年)。雄性和雌性海龟异步迁移,与男性提前到达繁殖地点周雌性(Limpus 1985;多德1988)。然而,它是可能的,一些男性non-migratory,筑巢海滩附近,保持全年(Henwood撰写的1987)。
龟,展品主要是近岸迁移的另一个物种是平背海龟(游泳者depressus)。这些海龟最限制迁徙所有海龟的范围。他们喂养区印尼和新几内亚,迁移和高度的忠诚在澳大利亚北部海岸筑巢海滩每1 - 3年(Limpus et al . 1981 1983 1984;Parmenter 1994)。Postnesting迁移可能数百到数千公里的长度,和一般限制在澳大利亚北部近岸水域,至于巴布新几内亚(西方)。
棱皮龟(Dermochelys coriacea)展示一个完全不同的物种的迁徙模式的海龟上面所讨论的。这个物种高度游牧,进行越洋迁移,并能潜水,伟大的深度(高夫et al . 1994;Morreale et al . 1996;埃克特1998;海斯,霍顿et al . 2004年)。棱皮龟不利用固定捕食场所(海斯,霍顿和迈尔斯2004年,但北大西洋的一些地区可能会定期访问捕食场所);相反,他们迁移网站之间广泛(Keinath Musick和1993;Steyermark et al . 1996;海斯,霍顿et al . 2004年)(图4 c),并经常发现在寒冷的北部海域远离筑巢区(Pritchard 1976;海斯,霍顿和迈尔斯2004)。 The pre-nesting migrations of leatherback turtles are not well understood; however, females are known to migrate to nearshore waters adjacent to nesting beaches a few weeks prior to the nesting season every 2-3 years. Unlike other marine turtles, female leatherbacks do not demonstrate strong nest site fidelity, although they do return to the same general nesting area. After nesting, females migrate extremely long distances, sometimes across oceanic basins (Morreale et al. 1996; Eckert 1998; Hughes et al. 1998). It is not known, however, whether or not these long distance movements follow generalized tracks. Indeed, major oceanographic processes, such as main currents and eddies, have been shown to have a major influence on Leatherback movements, placing into question whether these journeys are really migrations or just prolonged stays in vast feeding areas (Luschi et al. 2003b).
棱皮龟、榄蠵龟龟(摘要本文报道)高度洄游(Polovina et al . 2004年),和花大部分的非生殖生活在开放海域(海狸和卡萨诺1996;普罗金2003)。在筑巢季节繁殖活跃的男性和女性迁移向海岸,和总在筑巢海滩附近的繁殖地。女性出现在海滩上产卵,尽管他们并没有表现出很强的巢穴富达(卡尔布1999)。一些男性这个物种似乎non-migratory,沿着迁徙路线,而是试图拦截雌性筑巢海滩,因此不同的男性利用不同的生殖策略(普罗金et al . 1995;Kopitsky et al . 2000年)。交配后,女性仍然在繁殖地区几个星期甚至几个月。榄蠵龟龟的post-nesting迁移是独特的和复杂的。采取的路线雌性个体和群体的女性每年不同,和似乎并不遵循任何形式的迁徙走廊(普罗金2003)。榄蠵龟的高度游牧,遍历一望无垠的大海没有利用任何特定的,固定的捕食场所。
总的来说,海龟证明三个不同的迁徙模式(图4)。一些物种,包括大海龟和榄蠵龟龟,没有一个固定的觅食区,在广泛的和不可预知的回到他们的繁殖区域(图4 c)。相反,坎皮海龟,争执不休,背龟繁殖地区之间的迁移和高产的浅海大陆架上觅食区域(图4 b)。最后,绿海龟和玳瑁海龟利用良好,固定觅食区域,很少的变化(图4)。叠加在这个较小的迁徙运动由海龟在季节性的基础上他们前往温暖的觅食为由冬季时期(Gitschlag 1996)。
海龟迁徙的许多方面仍然不确定。例如,研究主要集中在postnesting迁移,因为这些更容易比prenesting迁移研究。此外,男性海龟的运动不是很好理解,因为他们不上岸,因此很难标签。有限radiotracking男性的研究表明,每年他们忠实地回到同样的育种领域(菲茨西蒙斯et al . 1997年)。然而,其他研究表明,一些物种的雄性可以non-migratory (Henwood撰写的1987;普罗金et al . 1994;Kopitsky et al . 2000年)。最后,小海龟孵化后的运动模式不是很了解(图5)。人们认为幼仔的一些物种漂移旋转,或圆形电流系统,作为移动,大洋苗圃场地(Luschi,休斯et al . 2003年)。小海龟在大西洋,因此逐渐迁移一个往返,12年,并最终回到自己的出生地的海滩繁殖(海尔斯顿1994;罗曼et al . 2001; Pough et al. 2002). Juveniles and subadults of many populations reside in coastal feeding areas located hundreds or thousands of kilometers from their natal beaches (Fig. 5). For example, juvenile Loggerhead turtles from nesting beaches in Japan and Australia traverse the entire Pacific Ocean (Bowen et al. 1995), and young Loggerheads from nesting beaches in the western Atlantic will traverse the entire Atlantic Ocean in the course of their developmental migrations (Bolten et al. 1998).
一些其他类型的龟也进行嵌套迁移,尽管他们不是广泛的海龟。例如,欧洲陆龟(陆龟hermanni) (Swingland et al . 1986年),和一些淡水龟,如Glyptemys insculpta(奎因和泰特1991),Kinosternon(长臂猿et al . 1990), Chelydra serpentina(长臂猿et al . 1990年),和Chelodina longicollis (Graham et al . 1996年)被发现进行季节性迁徙的50米到5公里从池塘或森林地区筑巢地点。一些物种的雌性表现出强烈的忠诚一般筑巢区,和将返回相同的水生栖息地年复一年
(林德曼1992;多德2001)。然而,海龟也被观察到具有迁移的原因除了找到合适的筑巢地点。例如,鳄龟(Chelydra serpentina)可能迁移到4公里超出其正常活动范围寻找合适的过冬地点在流水(礼物1944;布朗和布鲁克斯1994)。亚巨型陆龟(Geochelone gigantea)从内陆地区迁移到沿海地区在雨季获得高质量的食物资源在海岸(Swingland和Lessells 1974;Swingland et al . 1989年)。加拉帕戈斯陆龟也不同栖息地之间的迁移(Rodhouse et al . 1975年)。许多淡水龟已经观察到迁移到新池塘由于波动的环境条件等干旱(贝内特et al . 1970;美国约曼公司1995;格雷厄姆et al . 1996年)。然而,在两栖动物中,有某种程度的变异在种群和物种之间的迁徙模式的海龟。例如,如果合适的筑巢地点附近是可用的,欧洲陆龟不会迁移(Swingland et al . 1986年)。此外,迁移可能是危险的,大力昂贵,或者两者都是,只有大约20%的亚乌龟人口研究迁移到海岸利用丰富的食物资源(Swingland et al . 1989年)。
除了海龟,一些鳄鱼也进行定期、季节性迁移。一些物种,包括尼罗河鳄鱼(Crocodylus niloticus)可能游泳很远的地方寻找合适的嵌套网站(Chelazzi 1992)。没有观察到这个物种的雌性繁殖中央岛上的海滩游泳,鲁道夫湖产卵Modha (1967 1968)。然而,这种行为在鳄鱼是罕见的,和大多数物种,包括澳大利亚Crocodylus johnstoni,仍在本国范围内嵌套(韦伯et al . 1983 a, b)。一些种类的鳄鱼,鳄鱼在繁殖季节可能会增加他们的活动范围,但是像鳄鱼一样,筑巢地点通常位于活动范围,和动物没有进行嵌套迁移(Pough et al . 1998年)。此外,尽管一些鳄鱼旅行很长距离,例如跳岛游的行为表现出通过盐水鳄鱼,这些运动不考虑迁移(瑞达1985)。在一些鳄鱼,迁移,以应对环境变化比嵌套迁移更常见。一些种类的鳄鱼(Crocodylus johnstoni),凯门鳄(凯门鳄crocodilus),从干燥沼泽和短吻鳄季节性迁移到更永久的水域(尼尔1971;Pough et al . 1998年)。
-
 emerging from a communal den in the spring in the Interlakes region of Manitoba, Canada. Photograph courtesy of Dr. Herb Rosenberg")
- 图6所示。大量积累的一部分red-sided花纹蛇(Thamnophis sirtalis parietalis)从公共巢穴在春天Interlakes地区的马尼托巴省,加拿大。照片由草罗森博格博士
 emerging from a communal den in the spring in the Interlakes region of Manitoba, Canada. Photograph courtesy of Dr. Herb Rosenberg")
大多数卵生的有鳞的产卵在正常范围,因此不进行育种迁移,尽管一些种类的蛇已报告做定向运动专业产卵或繁殖地点(Pough et al . 2002年)例如,Vipera贝鲁在芬兰从姥网站常见的养殖面积几百米远(Pough et al . 2002年)。许多种类的蛇也表现出戏剧性的夏季喂养范围和冬季丹宁网站之间迁移。温带北部的蛇,这是很常见的,包括花纹蛇(Thamnophis)(图6)和响尾蛇(例如Crotalus atrox) (Landreth 1973;Klauber 1982;劳森1989;罗森和Secoy 1991;鲍尔和罗素2001;LeMaster et al . 2001;林基2001)。这些蛇一般对1-10km迁移到他们的窝点在秋天冬天冬眠。 Snakes congregate at the denning sites, and mating occurs at the dens before the snakes migrate back to their summer feeding areas in the spring (Klauber 1982; Lawson 1989; LeMaster et al. 2001) (Fig. 6), although the patterns of seasonal movement may differ between males and females (King and Duvall 1990). Like other migrating reptiles and amphibians, garter snakes and rattlesnakes tend to show fidelity to both their den sites and their feeding areas, returning to the same locations year after year (Lawson 1989). The sea snake, Pelamis platurus, also undertakes long distance breeding migrations in the Indian and Pacific oceans (Graham et al. 1971).
其他类型的蛇展示不同的迁徙模式。例如,水蟒(Liasis fuscus)在澳大利亚每年迁移距离相当远跟着他们最重要的食物来源,忧郁的老鼠(鼠colletti)。尽管许多大型哺乳动物迁移到跟随猎物,这种类型的迁移陆地冷血动物是很困难的,因为它们通常小,持续活动的能力有限,往往不活跃今年大部分时间由于热约束(马德森和发光1996)。水蟒是大型动物,生活在热带环境中,因此能够承担长途迁移追踪猎物。这些蛇已经观察到遵循他们的主要猎物12公里backswamp地区泛滥平原的雨季(马德森和发光1996)。其他类型的水生蛇,像阿拉弗拉filesnakes (Acrochordus arafurae),也进行季节性迁徙,从限制池塘草原淹没在雨季的开始。因此表现出强烈的季节性变化的栖息地利用的可能是由于猎物后的蛇(鱼)的池塘,或可能只是由于环境条件的变化(光泽和兰贝克1985)。
在爬行动物中,迁移是最普遍的蜥蜴。这些陆生动物经常显示强劲,领土附件到特定的网站,他们的体积小,冷血动物自然倾向于排除迁移(马德森和发光1996)。然而,一些种类的蜥蜴已经观察到的迁移。大型iguanids生活在环境中嵌套稀缺的网站可能会迁移到15公里到达合适的筑巢地点(Pough et al . 1998年)。例如,绿鬣蜥Barro科罗拉多岛游到小离岛巢,尽管大多数移动小于1公里超出其正常活动范围(兰德1968;一杯啤酒et al . 1985年)。类似的模式出现在女性地面鬣蜥(Cyclura spp),迁移至6.5公里到达合适的筑巢地点(Wiewandt 1982)。加拉帕戈斯群岛陆地鬣蜥(Conolophus subcristatus)也迁移到筑巢地点,和一些女性已经观察到爬1400火山火山口的边缘,然后下降900米到火山口巢(Werner 1983)。
有些不寻常的迁徙行为也被观察到在西澳大利亚Ctenophorus蜥蜴。青少年移民远离岩石露头2 - 3个月大时避免骚扰的成年男性。一旦青少年达到性成熟他们迁移回到主露头(布拉德肖1971)。大多数其他蜥蜴不迁移,但许多lacertids和iguanids展览自导能力(梅休1963;温特劳布1970)。
继续阅读:导航和定位的爬行动物
这篇文章有用吗?
读者的问题
-
艾登5个月前
- 回复
-
亚伦6个月前
- 回复
-
1月6个月前
- 回复