导航和定位的爬行动物
导航和定位在爬行动物的迁移,扮演非常重要的角色,尤其是在长途迁徙的海龟。海龟的生活包含了一系列的迁移从他们出生的幼仔开始游泳沙滩大海,然后成年海龟觅食区域之间的迁移和筑巢海滩(图5)。这些动物长途迁徙在公海地标在很大程度上是不可用,但他们证明非常精确的定位到特定的位置,包括一些非常小的岛屿(楚格et al . 2001年)。海龟的生活方式从而密不可分的能力东方跨跨度似乎毫无特色的海洋准确,表明这些动物拥有一个复杂的定位系统。
视觉线索
海滩
区
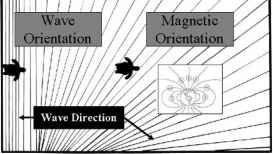
图7所示。顺序方向线索猜测指导人工孵化的海龟筑巢海滩到开放的海洋。修改后,罗曼et al。(1997年,图5.8)
人工孵化的海龟的迁徙运动同样令人印象深刻的比成年人(图7)。这些小动物爬向大海,并立即建立一个离岸航向保持很久之后的陆地已经丢失。孵化后,年轻的海龟使用三种不同类型的线索来东方(Pough et al . 2002年)(图7)。首先,在新兴的巢,小海龟利用视觉线索,东方走向大海。研究表明,人工孵化的海龟和绿海龟
视觉线索
开放的海洋
波折射
开放海域表现出强烈的phototropotactic反应,将东方最亮的光在附近,,在没有人工照明的情况下,所反映出的地平线在海洋,海面反射的光(卡尔1962;金斯米尔和Mrosovsky 1982;Mrosovsky和Kingsmill 1985;鲑鱼和Wyneken 1990;彼得斯和Verhoeven 1994)(图7)。使用的颜色或波长的光也是人工孵化的海龟,帮助他们找到大海。出于这个原因,人工照明附近海滩幼仔可能是一个问题,导致他们变得迷失方向和扰乱sea-finding能力(鲑鱼2003)。绿海龟表现出偏爱蓝色或紫色的波长在红、橙色或黄色光(Witherington和Bjorndal 1991;罗曼et al . 1997年)。这种偏好使他们避免定向向上升或落日。偏振光也可以使用幼仔东方向海。最后,幼仔倾向于避免黑暗,轮廓升高,这可能是由于沙丘或植被,对象通常位于一个方向相反的海岸线(Godfrey 1995;罗曼et al . 1997年)。这种行为也有助于指导幼仔向海,和类似的线索可以使用嵌套成人后重返大海产卵(考德威尔和考德威尔1962)。
一旦他们到达海洋,大力幼仔开始游泳,由波暗潮5-10m从海滩上完成。表面时,小海龟建立课程走向大洋,用波传播方向东方(图7)。他们游泳垂直于海浪,导致他们进入海洋,向全球当前系统(鲑鱼和罗曼1989;罗曼et al . 1990 1995;罗曼1992;Pough et al . 2002;Nagelkirken et al . 2003年)。
然而,远离海岸,波方向不再提供了一个可靠的海上方向指示的,然而幼仔继续遵循相同的外海。此外,幼仔进行数千公里从他们出生的海滩回到品种,有时30 - 50年。这表明他们使用其他更复杂的来源方向信息。研究表明,人工孵化的海龟和棱皮海龟对地球磁场敏感,有人建议他们开发一个磁罗经离开他们出生的海滩(罗曼1992;罗曼et al . 1997;欧文et al . 2004年)(图7)。例如,幼仔训练游泳在一个特定的方向,朝着光线会保持同样的指南针方向甚至在黑暗中。此外,如果磁场逆转,海龟会扭转他们游泳方向(罗曼et al . 1997年)。
小海龟在海里也显示能力之外的一个简单的对地球的磁场。有证据表明他们有能力探测倾角的变化的地球磁场相对于地球的引力场(光et al . 1993年),以及当地磁场强度的变化。例如,人工孵化的海龟没有迁徙的经验已经证明东方相对于磁场的方向,这样他们就能在温暖的北大西洋环流目前的系统,在一个安全的迁徙路线(罗曼和罗曼1998;罗曼et al . 2001年)。的能力来检测倾角和地球磁场的强度将允许海龟近似全局位置的纬度和经度,向他们提供一种bi-coordinate地磁图(罗曼et al . 1997年)。
这些幼仔的研究可能提供洞察导航提示受雇于成年海龟,虽然有可能是青少年和成人应对不同类型的线索。迁移的成年海龟一直得到广泛的研究在过去的十年,但这些动物使用的定位和导航机制仍然是不确定的(爸爸和Luschi 1996;爸爸et al . 2000年),尽管它可能涉及多种感官刺激,两栖动物一样(海斯et al . 2002;Pough et al . 2002年)。甚至有一些争论这些动物是否能够真正的导航。有人建议,因为他们遵循他们的目标相当直的路线,简单的罗盘定位是足够的指导他们的迁移(Luschi,休斯et al . 2003年)。这可能是适用于迁移对大目标,如绿海龟的返回迁移到巴西海岸(图4),但可能不是嵌套迁移,这是指向一个很小的目标。流离失所的成年海龟在印度洋被发现无法弥补post-nesting位移和似乎依赖于简单的定位机制,如海岸线,作为指导(Luschi,海斯和爸爸2003)。嵌套迁移要复杂得多,可能需要动物mapsense,和真正的导航能力(爸爸et al . 2000;Hays et al . 2002年)。 Furthermore, the ability of displaced turtles to return to or orient towards their original location (Papi and Luschi 1996; Luschi et al. 1998; Avens and Lohmann 2004), and the ability of turtles to correct deviations in their migratory course caused by ocean currents (Luschi et al. 1998; Papi et al. 2000), suggest that simple compass orientation is not adequate, and that a more sophisticated mechanism is being used.
如果检测磁场的能力已被确认的幼仔被保留在成人中,那么很可能成年海龟也使用磁定位和导航地图(Lutcavage 1996;罗曼et al . 1997;爸爸et al . 2000;罗曼et al . 2004年)。然而,如果存在这样一种能力,它还不知道如果它只提供非常精确或近似natal海滩的信息,与其他类型的信号使用接近出生的海滩。此外,追踪绿海龟的迁徙路线的一项研究发现,磁干扰采取的路线海龟的控制海龟没有显著差异,表明磁信号不是必不可少的海龟的迁移(爸爸et al . 2000年)。另一方面,本研究关注的返回迁移海龟,海龟,从而有可能更多地依赖于磁信号同时进行更复杂的pre-nesting迁移(爸爸et al . 2000年)。
除了磁信号,提出了许多其他机制来解释海龟的定向能力。例如,在两栖动物中,嗅觉可能发挥重要作用在这些动物的迁移(卡尔1967;曼顿et al . 1972;鲍恩et al . 1989;Chelazzi 1992;爸爸et al . 2000年)。卡尔(1967)发现证据表明,海龟有至少一年的气味记忆,导致其他化学线索表明青少年海龟印记与出生的海滩,然后使用一种化学羽洋流,加上指南针方向,指导他们海滩当他们返回作为成年人(罗曼et al . 1997;爸爸et al . 2000年)。然而,化学信号在远距离导航的作用几乎是未知的,也没有令人信服的实验证据支持这一假设。此外,直接迁徙路线的成年人是不一致的与跟踪化学羽,和海洋洋流信风带来的可能会扰乱化学线索,从而防止化学的形成轨迹,海龟可以效仿(1990年布朗)。它也是值得商榷的曝光时间natal海滩的气味是足够长的时间的印迹幼仔。基et al .(2003)审查相关证据的方法绿海龟(龟鳖目mydas)可以使用定位阿森松岛的迁移从巴西海岸。调查取代女性的行为,他们得出的结论是,化学信息经由洋流没有导航,占bi-coordinate地磁方法,也没有其他磁梯度或天体的线索。相反,女性发现岛上搜索和报警,然后靠风传播的信号用于最终位置。
另一个可能性是海龟东方使用波方向或洋流,观察在人工孵化的红海龟,他们这个系统校准使用天体线索如星星和太阳(Morreale et al . 1996;爸爸et al . 2000年)。其他潜在的导航机制包括信息从海面上的风(Luschi et al . 2001;基米et al . 2003;Hays et al . 2003),海底水深的特性(Morreale et al . 1994年),或水温(普罗金2003)。然而,稀疏的证据来支持这些机制,一些已经被证明在一些物种的迁移不重要的海龟。例如,绿海龟不依赖海洋表面温度而迁移(海斯et al . 2001年)。总体而言,更多的研究是必要的任何可以得出明确的结论之前关于海龟的定位和导航系统。
除了海龟,取向和归巢机制也被其他类型的爬行动物包括淡水和陆地龟,蛇,鳄鱼和一些蜥蜴。虽然一些种类的淡水和陆地龟进行广泛的迁移,已发现许多拥有令人印象深刻的导航能力。例如,地图龟(Graptemys pulchra)回家后流离失所的24公里沿着一条河,这让他们远远超出了自然活动范围的边界(谢伊1976)。然而,海龟的总数,返回成功很低。许多其他淡水龟类物种,包括Clemmys guttata, Chrysemys picta, Glyptemys insculpta和Apalone mutica,已被证明具备良好的导航性能后位移(礼物1944;安永1968 1970;普卢默和夏勒1975;卡罗尔和艾伦菲尔德1978)。淡水龟东方的机制还不是很清楚。实验表明,海龟的定向能力属Terrapene和Chrysemys减少阴天也是如此,尽管一些仍然能够回家短距离多云的天空(Emlen 1969; Yeomans 1995; Graham et al. 1996). These results indicate that these turtles orient using a sun compass calibrated with an internal clock (Gould 1957; DeRosa and Taylor 1980 1982; Yeomans 1995; Graham et al. 1996). However, olfactory cues may also play an important role in homing and orientation of freshwater turtles. Olfaction has been suggested as an important factor in the homing ability of Glyptemys insculpta (Carroll and Ehrenfeld 1978), and olfaction and geotaxis could not be ruled out as factors in the homing of Chrysemys picta (Ernst 1970). Furthermore, the Australian Snake-necked Turtle, Chelodina longicollis, has been found to orient towards its home substrate in laboratory experiments (Graham et al. 1996), suggesting that olfaction also plays a key role in the orientation of these animals. It has furthermore been suggested that some freshwater turtles have the ability to detect and orient towards water, indicating that they may be using surface-reflected polarized light to orient, as polarization tends to be higher over moist surfaces (Yeomans 1995; Zug et al. 2001). Finally, the species Terrapene carolina has been shown to use both local visual cues (Lemkau 1970) and magnetic cues to aid in orientation (Mathis and Moore 1988).
乌龟的导航和定位机制不太理解。乌龟陆龟hermanni能够定位在1560米外的距离其活动范围(Calzolai和Chelazzi 1981;Chelazzi和Calzolai 1986)。有人建议,乌龟用嗅觉线索向本国范围,东方和定位后水体的位移(Chelazzi和参与1986;Pough et al . 2002年)。乌龟也可以利用视觉线索基于地形起伏(Auffenberg和韦弗1969;McCoy et al . 1993),但在大多数情况下没有明确结论乌龟取向。
导航行为鳄鱼中也得到了证实。其中最令人印象深刻的能力一直在观察青少年短吻鳄。一般来说,鳄鱼是相当久坐不动的动物,但会迁移,以避免不利的环境条件。研究表明,鳄鱼能够回家从5公里远离他们的捕获网站,并能够对从东方到34公里(瑞达1985)。像海龟,鳄鱼可以检测磁场,因此他们使用磁场取向(瑞达1984)。此外,他们能够从陌生的区域,这表明他们有能力真正的导航(瑞达1984 1985)。其他种类的鳄鱼,包括凯门鳄crocodilus和咸水鳄鱼(Crocodylus porosus)还发现了回家后位移分别为2公里和30公里(Gorzula 1978;Ouboter和Nanhoe 1988;Pough et al . 2002年)。
自导能力在一些蛇似乎也常见,但结果有关的北美蛇一直矛盾的。导航行为已经观察到在若干种花纹蛇(Thamnophis sirtalis, t .基数),水蛇(Nerodia sipedon, Regina septemvittata)和响尾蛇(Crotalus atrox)当他们流离失所的从他们的巢穴地点或油腻的食物来源(Landreth 1973;新人et al . 1974;布朗和帕克1976;劳森1989 1994;劳森和Secoy 1991)。然而,花纹蛇没有演示了相同的证据向家的方向当流离失所的正常范围(1994年Lawson)。其他种类的蛇,包括Sisturus catenatus和Carphophis享乐也不表现出导航行为,并将建立新的家庭范围当流离失所(Barbour et al . 1969;看看和Kodrich 1982)。基于这些结果可能有些蛇可以导航的能力,但他们可能并不总是愿意使用这些能力。
一些蛇,做展览归航行为能够沿着特定的东方指南针方向,即使流离失所的从他们的活动范围或巢穴地点很远(Landreth 1973;新人et al . 1974;布朗和帕克1976;劳森1989 1994;劳森和Secoy 1991)。此外,研究表明,取向方向上被蛇变化如果它们在黑暗中运输,这表明这些动物使用天体暗示取向(新人et al . 1974;格里高利et al . 1987;劳森1989;劳森和Secoy 1991)。使用何种类型的天体提示可能是太阳能,因为没有证据表明夜间蛇利用月球或恒星线索取向(Chelazzi 1992)。 Finally, there is some evidence to suggest that some snake species use olfactory cues for short-distance orientation (Diadophis punctatus: Dundee and Miller 1968), and that juvenile Thamnophis and Crotalus follow the pheromone trails of conspecifics to guide them towards their goals (Ford 1986; Graves et al. 1986; Lawson and Secoy 1991; Ford and Burghardt 1993; Pough et al. 1998).
蜥蜴,迁徙行为是非常罕见的,完全不知道他们的导航和定位。几乎只在lacertids自导能力蜥蜴被发现和iguanids(1963年5月凿;温特劳布1970)。几个种Sceloporus,由于和Uta能够归航后位移小于300从正常范围(Spoekker 1967;盖伊1978;Bissinger 1983;Ellis-Quinn和西蒙1989;Chelazzi 1992)。其他蜥蜴物种已经证明的能力回国后短位移(Dipsosaurus背的:Krekorian 1977;Takydromus takydromoides:石原1969; Podarcis sicula: Foà et al. 1990; Tilqua rugosa: Freake 1998). However, many other lizards are not capable of homing from similar distances (Anolis lineatopus: Rand 1967; Sceloporus occidentalis: Fitch 1940; Uta stansburiana: Tinkle 1967). It has been suggested that the strength of attachment to a home range affects the homing ability of lizards. For example, male and female Sceloporus jarrovii are both territorial, and demonstrate similar homing abilities. Conversely, nonterritorial female Sceloporus orcutti exhibit poorer homing abilities than territorial conspecific males (Ellis-Quinn and Simon 1989; Weintraub 1970). Sex-based differences in homing behavior have also been observed in Tilqua rugosa (Freake 1998). Similarly, the territorial species Sceloporus graciosus has better homing abilities than the non-territorial species Phrynosoma douglasii, even though they occupy the same habitat (Guyer 1991).
取向受雇于蜥蜴的机理还不是很清楚。实验使用定向训练蜥蜴建议使用天体罗盘在蝎虎星座viridis菲舍尔(1961)、乌玛notata(阿德勒和菲利普斯1985)和Tiliqua玫瑰(Freake 1999)。蜥蜴的松果腺器官可能充当偏振光探测器,因此一些蜥蜴可能能够使用偏振光,东方蝾螈的一些物种一样(Ellis-Quinn和西蒙1991;Freake 1999;楚格et al . 2001年)。然而,尽管许多蜥蜴物种能够导航和定位,大多数不进行迁移,这些能力可能有用的其他一些功能在这些动物。
继续阅读:相关的人口结构模式钝口螈属macrodactylum在空间和时间
这篇文章有用吗?