获得综合的理解棕囊藻属spp Prymnesiophyceae通过modeldriven实验室和mesocosm但是此属的研究
斯图尔特·j·惠普尔•伯纳德·c·彭定康•彼得·g·维里蒂•马克·e·弗里希•杰里米·d·长•Jens c . Nejstgaard•乔恩·t·安德森•安尼塔·雅各布森•Aud拉森•华金Martinez-Martinez•斯图亚特·r·Borrett
收到:2006年4月20日/接受:2006年9月7日/网上公布:2007年3月8日©Springer科学+商业媒体帐面价值2007
抽象的知识的复杂的生命周期棕囊藻属是一个关键的理解但是此属它在海洋生态系统中的作用和全球生物地球化学。现有的生命周期模型的修改和用于集成理解棕囊藻属。但是此属
美国j·惠普尔•p·g .真实性•m·e·弗里希Skidaway海洋学研究所,萨凡纳,美国佐治亚州
佐治亚大学生态研究所,雅典,
30602年乔治亚州,美国
电子邮件:(电子邮件保护)
Nahant海洋科学中心,东北大学,硕士,美国
j . c . Nejstgaard•a . Jacobsen•a . Larsen生物学系,卑尔根,挪威卑尔根大学
j·t·安德森
河口研究中心,摩根州立大学圣伦纳德,医学博士,美国
j . Martinez-Martinez普利茅斯海洋实验室,英国德文郡普利茅斯
计算学习实验室,斯坦福大学,斯坦福大学,加州,美国
生命周期。在模型驱动的研究中,模型暴露我们的理解不足,实证研究接踵而至,和反馈改进的理解。这个方案后,生命周期模型的三个方面进行。有四个例外,描述的实证研究提出了其他文献引用。第一个方面涉及测试存在的一个过程或产生它的描述。这些研究包括:体外集落部门演示棕囊藻属pouchetii但是此属、描述的体外p . pouchetii的殖民地形状变化与衰老有关,确定哪些p pouchetii人生阶段很容易受到病毒感染和细胞溶解,并设计一个实验来确定新棕囊藻属殖民地,但是此属的沉积物可能是来源上覆水域;结果表明,底栖生物粒子的更细节的调查作为群体形成的物理衬底是十分必要的。第二方面涉及的调查过程率量化或过程控制参数。流程率量化殖民地部门包括测量率和增长率使用mesocosm-derived殖民地。测试过程控制实验包括硅藻细胞膜增强p pouchetii从孤独的细胞集落形成,和调查mesozooplanktonic抑制和microzooplanktonic增强棕囊藻属globosa但是此属集落形成浮游食草动物的信息物质。第三方面开门的分子识别单个细胞之间的基因差异,p . globosa殖民地。这些研究旨在提供洞察问题的控制因素参与单细胞和殖民生活之间的过渡阶段。生命周期模型提供了一个现成的地方新见解和理解从实证研究合并到现有的模型,并且可以用于提高仿真模型的直接和间接影响棕囊藻属全球生物地球化学。但是此属
关键词概念模型研究•••Mesocosm•生命周期模型驱动的棕囊藻属。但是此属
介绍
属棕囊藻属包含物种,但是此属可能扮演了一个重要的角色在全球生物地球化学(Jr史密斯et al . 1991;兰斯洛特et al . 1994;Wass-mann et al . 2005年)。他们出现在广泛的领域(Stefansson和Olafsson 1991;兰斯洛特和卢梭1994),供应的重要部分全球二甲基硫(DMS);Gabric et al . 1999;艾尔斯和吉列2000)和浮游生态系统中扮演着重要角色(真实性和斯1996;兰斯洛特et al . 1998年)。生命周期过程的棕囊藻属可能对生态系统生物地球化学过程有显著的影响包括但是此属营养吸收和再生生命形式的棕囊藻属,但是此属营养释放由病毒裂解,养分释放与浮游动物放牧、衰老和养分释放的殖民地。的细节知识的当前状态的棕囊藻属生命周期中全面回顾了但是此属卢梭et al。(在媒体,这卷)。 In order to integrate understanding of the Phaeocystis life cycle, a conceptual model was constructed (Whipple et al. 2005a), although the existence of some life stages was uncertain and the controls on many of the processes were poorly understood. This exercise highlighted experiments which could provide a better understanding of the role of the life cycle in species dynamics和生态系统过程。
这种增强的理解将被纳入发展概念模型(Whip-ple et al . 2005年),并到一个生态系统模型的棕囊藻属生命周期只有一个但是此属的生物隔间(惠普尔等人unpubl)。在模型驱动的研究中,模型展示在我们的理解Phaeo-cystis生命周期内嵌在它的生态系统;实证研究来解决这些漏洞接踵而来,这个循环的反馈迭代改进的理解。本文例子的生命周期模型,棕囊藻属被用来直接但是此属研究棕囊藻属生命周期及其生态,但是此属和演示了如何洞察这些研究集成到一个概念模型上下文提高和集成这样的实证结果。
模型描述
被修改的棕囊藻属生命周期,但是此属从模型提出了惠普尔et al .(2005),进展顺时针方向从左上角图1与二倍体的细胞,作为起点。这些细胞流与新的殖民地,是孤独和殖民之间的重要过渡阶段的生命形式。
新的殖民地细胞第一次分裂后,他们转移到越来越多的小殖民地隔间。成熟继续通过大小类:小!媒介!大细胞通过有丝分裂和粘液的殖民地。还有backtransfer通过菌落大小从大到小类划分(惠普尔et al . 2005 b)。
当殖民地开始失去细胞,种植殖民地转移到基于相应大小的衰老殖民地。二倍体或单倍体单独细胞释放这些完成棕囊藻属生命周期。但是此属的四个克隆形成棕囊藻属的物种,但是此属单倍体鞭毛虫只记载棕囊藻属globosa但是此属(Eilert-sen 1989;雅各布森2002;Medlin和Zingone出版社,这卷;卢梭等人的新闻,这卷)。其他生命周期输入二倍体的细胞来自于转换的新殖民地。两性生殖并没有被记录在任何棕囊藻属的物种,但是此属不过,它已经包含在创建所需的模型,因为它假设为二倍体的细胞从p . globosa hap-loid细胞集落形成之前(卢梭等人在媒体,这卷)。因为这
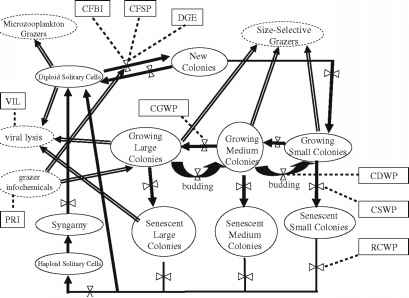
图1棕囊藻属生命周期图。但是此属实线椭圆:棕囊藻属生命周期阶段但是此属;虚线椭圆:其他生态系统组件。实心箭头:生命周期转换;波谷线箭头:输入或输出从棕囊藻属。但是此属[X]
符号:生命周期转换速率控制。与虚线矩形缩写:实证研究指向一个相关的生命周期阶段或生命周期过渡
图1棕囊藻属生命周期图。但是此属实线椭圆:棕囊藻属生命周期阶段但是此属;虚线椭圆:其他生态系统组件。实心箭头:生命周期转换;波谷线箭头:输入或输出从棕囊藻属。但是此属[X]
符号:生命周期转换速率控制。与虚线矩形缩写:实证研究指向一个相关的生命周期阶段或生命周期过渡
实验描述的名称和缩写:
CDWP殖民地部门在孔板
经验与种子集落形成颗粒
CFBI集落形成与底栖生物的参与
CGWP殖民地孔板的增长
在孔板CSWP殖民地衰老
DGE差异基因表达
革命制度党棕囊藻属应对信息物质。但是此属
RCWP孔板细胞释放殖民地
维尔病毒感染和裂解模型开发代表属棕囊藻属,但是此属包括单倍体的生活阶段,但可能只发生在某些物种(惠普尔et al . 2005年)。
波谷线箭头在图1表示假设流或从non-Phaeocystis组件更大的生态系统。在某些情况下,这些作为对生命周期的控制流,和在其他情况下边界输入或输出的棕囊藻属生命周期。但是此属三个主要的输出从棕囊藻属生命周期表示。但是此属两个放牧在孤独的细胞(左上),和放牧在所有大小类的殖民地(右上)。第三是病毒裂解(左中),这是一个输出的过程通常强调在病毒的研究。然而,感染,一个输入过程,必须发生病毒来完成他们的生命周期和影响棕囊藻属细胞。但是此属最后一个过程是输入的信息物质,浮游动物食草动物(左下)成棕囊藻属单独cell-to-colony转换或殖民地但是此属细胞。
案例研究
本文的概念模型有两个功能:(1)为集成提供一个上下文是什么,和(2)定义的观察和实验,以消除未知(Whip-ple et al . 2005年)。图1以图形方式描述了如何棕囊藻属生命周期模型和实证研究来解决但是此属未知可能是相互关联的;下面描述每个实验在我们研究小组用缩写
表1描述的棕囊藻属生命周期过程和与它们相关的研究但是此属本文中讨论
图1中使用缩写名称过程描述
CDWP殖民地部门在孔板
CFBI集落形成与底栖生物的参与
经验与种子集落形成颗粒
CGWP殖民地孔板的增长
在孔板CSWP殖民地衰老
DGE微分gene_expression
革命制度党棕囊藻属应对信息物质。但是此属
RCWP释放殖民地细胞孔板SIM物种识别的昌盛
维尔病毒感染和细胞溶解
(例如,维尔=病毒感染和细胞溶解;左中心)表明,它与生命周期的关系。9个流程及其相关研究认为本文所示图1的生命周期环境。这些过程的描述,他们的首字母缩写,数据来源如表1所示;一个条目描述验证的棕囊藻属物种但是此属mesocosm研究(SIM)也包括在内。阶段或过程描述的现象进行调查是与生命周期模型。括号缩略词表1的实验出现在本节的小标题。在四个案例研究,提出了详细的方法和结果;在其他情况下,一个简短的总结方法和结果将基于发布或提交文献来源,这可能是咨询详情。讨论考虑更广泛的影响的实证Wndings棕囊藻属生命周期动态本身,但是此属和潜在的生命周期模型的修正。
数据来源惠普尔et al . (2005 b)
Nejstgaard et al。(新闻)
惠普尔et al。(2005 b)本研究本研究
本研究本研究
雅各布森et al。(2005)
Mesocosm实验方法
许多下面讨论的观察和实验的上下文中进行mesocosm研究在挪威国家mesocosm Raunefjorden中心,位于海洋生物焊接站在西方挪威卑尔根大学的。研究进行了3月4日至2002年3月24日和2月27日,2003年4月2日(Nejstgaard et al . 2006);这些研究将被称为2002年和2003年mesocosm研究的其余部分。这些meso-cosm研究的主要结果将不会在这个手稿;他们提供详细Nejstg-aard et al。(2006)。本节的目的是提供足够的背景mesocosm研究,实验在他们的上下文可以被理解和解释。
在每个研究中,三个透明的聚乙烯外壳(深度为4.5米,2米直径,ca。11立方米,票面90%)充满了水过滤峡湾
在体外观察p . pouchetii殖民地部门
体外观察殖民地来自粗糙地面的材料收集的浅海湾沉积物体外观察p . pouchetii从孤独的细胞集落形成的硅藻细胞膜体外观察p . pouchetii体外观察菌落生长的菌落衰老基因差异表达p . pouchetii单独与殖民细胞p . globosa实验来确定p globosa应对微和mesozooplankton食草动物或其信息物质在体外观察集落细胞释放p . pouchetii 18 s rDNA调查评估棕囊藻属物种越来越多但是此属2003年挪威mesocosm体外观察病毒感染和裂解细胞完整和中断殖民地p . pouchetii从5米深度。一个空运系统完全混合enclosures每天大约五倍。以允许新物种的引入,避免pH值的变化,和更换水为抽样,10%的水在每个封装是每日更新峡湾(Nejstgaard et al . 2006年)。两三个meso-cosms修改与硝酸盐和磷酸盐与16的初始富集硝酸盐和1 | |米磷酸。每天添加硝酸盐和0.1 | 1.6 |米磷酸M取代营养被10%的水更新和抽样。在2002年的实验中,基于低养分浓度测量3月12日与额外的营养,营养丰富昌盛都增强硝酸8 | M和0.5 |磷酸M 3月12日晚。第三mesocosm在每个实验中,也许,作为控制(Nejstgaard et al . 2006年)。mesocosm设置的详细描述、采样、分析方法和结果描述Nejstgaard et al。(2006)。
物种鉴定昌盛(SIM)
在2002年和2003年mesocosm研究的许多棕囊藻属殖民地被观察到的是完美的球形,但是此属表明可能有p globosa而不是p pouchetii出现在西方的挪威峡湾。在第二次努力mesocosm实验(2003)确定的多样性棕囊藻属物种存在于这些mesocosm但是此属的研究。
方法
在整个2003年mesocosm研究、水(50500毫升)收集每隔两天到0.8 |米苏泊尔®膜过滤器(笼罩corp .)和存储在-80°C到分析。枚举的棕囊藻属sp.单一细胞和殖民地,但是此属两种样本选择分子棕囊藻属sp.但是此属的组成社区的决心。棕囊藻属sp.但是此属小亚基的核糖体RNA基因序列分析(rRNA)基因样本收集从受精外壳在实验初期只有单个细胞在场(2003年3月5日;单个细胞15 ml-1,殖民地0 ml-1)和样品收集开花高峰期间(2003年3月20日;单个细胞990 ml-1殖民地41 ml-1)检查。使用超净的总DNA提取过滤器™土壤DNA隔离(该款实验室,Inc .)如前所述(Allen et al . 2005年)。小亚基核糖体rna基因从棕囊藻属sp.但是此属特别聚合酶链反应(PCR)扩增使用通用18 s rRNA目标底漆UnivF-15 (5 ' ctg cca gta gtc ata tgc;弗里希et al . 2002年)和棕囊藻属genus-specific底漆PHAEO1但是此属(5 ' cgg tcg gg tgg cgt行动;兰格et al . 1996年)。放大反应是促进使用一个应用生物系统公司9700年20毫升thermocycler卷。 Each amplification reaction consisted of 8 ml Eppendorf ® MasterMix® (Eppendorf North America), 8.4 ml PCR grade distilled water, 3 ml each primer (100 ng ml-1), and 3 ml of undiluted or 1:10 diluted DNA extract. Purified DNA from the extracted samples was not quantified. Reaction conditions consisted of an initial denaturation step (94°C, 3 min) followed by 35 amplification cycles (94°C, 15 s; 53°C, 15 s; 72°C, 30 s) followed by a 7 min final extension at 72°C. PCR amplicons were examined on an agarose gel to insure that only a single band was produced. Single bands were cloned into the bacterial plasmid vector PCR®2.1 using the TOPO TA Cloning® kit (Invitrogen, Carlsbad, CA) and sequenced in both directions as previously described (Frischer et al. 2002).
结果
共有17个克隆测序和每个克隆序列的身份被证实是棕囊藻属pouchetii但是此属,表明它不太可能,任何其他种类的棕囊藻属,但是此属出现在这些研究。
测试存在的一个过程或生产过程的描述
集落形成与底栖生物的参与(CFBI)
底栖生物参与的生命周期不同的棕囊藻属物种已经讨论多年但是此属文学。凯塞(1970)描述了他的观察底栖生物细胞在文化如下:“有趣的是观察单个细胞的棕囊藻属pouchetii但是此属表现出倾向于附着于固体表面…我们可以假设在大海单一细胞附着在固体表面在海底或能动的粒子不断提供免费水与单个细胞或年轻的殖民地”(凯塞1970年,页209 - 210)。卢梭et al。(1994年,p . 26)得出结论,“……绝对没有一个真正的证据有区别的底栖生物阶段。”
卢梭et al。(在媒体,这卷)形容p . globosa二倍体鞭毛虫后一天内形成新的殖民地粘附到表面(Cariou et al . 1994;凯塞1970;Kornmann 1955;卢梭et al . 1994年)。非生物粒子(卢梭et al . 1994),文化血管壁(Cariou et al . 1994;凯塞1970)和硅藻(Weisse et al . 1986;Boalch 1987;卢梭et al . 1994年)已报告作为粘附网站(卢梭等人在媒体,这卷)。附件的细胞表面导致一个假设,即底栖生物阶段,作为过冬的形式,存在于自然环境(凯塞1970;卢梭等人的新闻,这卷)。 The general conclusion from these reports is that the diploid flagellate described in these studies is not a truly differentiated benthic form (Rousseau et al. in press, this volume).
实证研究在2003年mesocosm研究测试了p pouchetii单独细胞或殖民地将从峡湾resuspended沉积物孵化控制环境室的过滤水。
方法
底栖生物基质,由中等粒度的砂壳和生物遗体,被潜水员hand-collected从12米深度附近的沿海湾卑尔根,挪威在2003年3月14日。沉积物在隔离期间上覆水收集和实验准备、和孵化期间与过滤后的海水覆盖。一套20 cc整除的精疲力尽了,但潮湿的沉积物被安置在一式三份PC 250毫升瓶的底部,逐步覆盖200毫升以下水类型:人工海水;热压处理过的峡湾水;0.8 m ^过滤水mesocosm # 1(控制:没有营养补充);0.8 m ^过滤水mesocosm # 2 (NO3和PO4添加)。昏暗的灯光下的瓶子被孵化(1.7 m - 2 s - 1 ^摩尔,测量使用Biospherical仪器qsl - 100米的球形光收集器;十二12 L: D)在3°C步行在第0天冷的房间,采样(开始孵化)和第七天。的丰度、直径和p . pouchetii殖民地测量显微镜下的细胞密度。
结果
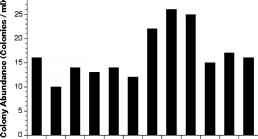
观察棕囊藻属pouchetii但是此属殖民地在湿地中治疗7天后孵化(图2)。最多的殖民地发展瓶包含过滤水从mesocosm 2 (M2)治疗,每天收到的氮和磷的增加(图2)。M2-water样本也包含了殖民地最大的直径,和最多的细胞每殖民地(无花果。2 b, c)。
p . pouchetii殖民地的存在部门(CDWP)使用的体外观察meso-cosm-derived p pouchetii殖民地在2002孔板,惠普尔et al . (2005 b)报道,1012%的水井包含至少一次分裂的殖民地。他们也观察到殖民地分工只是观察到许多小时后已经过去。这可能表明,殖民地经历一段时间的殖民地扩大通过细胞分裂和粘液生产之前分裂产生的女儿殖民地(惠普尔et al . 2005 b)。这种模式的殖民地乘法也报告p globosa(真实性et al . 1988年)。
标识区分单个细胞的分子标记和殖民地一阶存活率存在问题与各种形式的生命周期相关的棕囊藻属sp.但是此属是细胞的鉴定和遗传差异单和殖民细胞。开始解决这一问题的背景下复杂的棕囊藻属生命周期,但是此属差异显示PCR方法是用来识别基因的表达调控在殖民和单个细胞。这些研究进行了30
治疗
治疗b b
治疗
图2的结果收集地面的材料从浅海湾沉积物实验。治疗关键:ArtSW #:人工海水;峡湾AutoC #:热压处理过的水;M1 - #: 0.8 |米过滤水从mesocosm # 1(=没有营养补充);M2 - #: 0.8 |米过滤水从mesocosm # 2 (NO3和PO4添加)。(a)丰富的p . pouchetii殖民地(殖民地毫升”1);直方图代表殖民地估计为每个复制的瓶子。(b)菌落直径(|);误差线代表标准差的估计在每个瓶子从一数计算。(c)细胞密度(细胞殖民地”1); error bars represent standard deviations of estimates within each bottle calculated from triplicate counts
治疗
图2的结果收集地面的材料从浅海湾沉积物实验。治疗关键:ArtSW #:人工海水;峡湾AutoC #:热压处理过的水;M1 - #: 0.8 |米过滤水从mesocosm # 1(=没有营养补充);M2 - #: 0.8 |米过滤水从mesocosm # 2 (NO3和PO4添加)。(a)丰富的p . pouchetii殖民地(殖民地毫升”1);直方图代表殖民地估计为每个复制的瓶子。(b)菌落直径(|);误差线代表标准差的估计在每个瓶子从一数计算。(c)细胞密度(细胞殖民地”1); error bars represent standard deviations of estimates within each bottle calculated from triplicate counts laboratory cultures of P. globosa (CCMP 1528, Provasoli-Guillard National Center for Culture of Marine Phytoplankton, USA).
方法
三个独立的文化p globosa CCMP的1528人增长了一个星期在20°C,十四10:黑暗,L1-Si媒体(Guillard Har-graves, 1993)。在这些条件下单个细胞和殖民地都产生在每一个文化。单个细胞被反渗透分离殖民地10 |米孔筛和手动泵。细胞和殖民地分别收集到25毫米,5 |米Durapore过滤器(# SVLP02500,微孔,贝德福德,MA)。大约7.5 x 106单个细胞和5 x 104个小殖民地(ca。1毫米)固定化/过滤器。从每个示例使用RNA提取试剂盒RNeasy植物迷你包(猫# 74903,试剂盒,瓦伦西亚,CA)后,制造商的推荐程序(试剂盒,瓦伦西亚,CA)。纯化复制的RNA提取样本每个文化相结合(6抽取,每个细胞类型)。RNA被氟量计量化使用RiboGreen(分子探针,Inc .)和净化RNA是储存在“80°C到分析。
差异表达基因被确定微分显示ca的互补脱氧核糖核酸合成。1 | g总RNA的使用优势®cDNA PCR试剂盒(猫# K1905-1, Clontech实验室,Inc .,帕洛阿尔托,ca)。差异显示分析促进使用试剂和过程与商用三角洲微分显示装备®(猫# K1810-1, Clontech实验室,Inc .,帕洛阿尔托,CA)。放大的cDNA 90独特的组合来完成10任意25-mer引物(P引物)和9益生元(dT)引物(T引物)提供在三角洲地区的微分显示设备。微分显示放大反应进行了20 l |卷2 | l 10 x Klen Taq PCR反应缓冲区,14.2 | PCR年级蒸馏水,0.2 |混合核苷酸(5毫米),0.4 l | 50 x®优势KlenTaq, 0.2 | 33 p dATP, 1 | l 40或1:10稀释的cDNA模板。放大条件一样
表2差异表达基因片段p . globosa殖民和单个细胞
细胞类型。Cona部分监管完全监管乐队- - - - - -
君威Upa注册下来了
细胞1365 1193 32 32 117 16
殖民地1254 1193 7 7 7 15
一个案子,既定的表达;reg,调节;reg,表达下调Clontech推荐的,由三个low-stringency PCR周期(周期1:94°C, 5分钟;40°C, 5分钟;68°C, 5分钟),第二个和第三个周期(94°C, 30年代;40°C, 30年代;68°C, 5分钟),25附加high-stringency周期(94°C, 20年代;60°C, 30年代;68°C, 2分钟),后跟一个住在68°C周期7分钟。放大了在一个专用的增幅管thermocycler (Barnstead / Thermolyne迪比克,IA)。 Following confirmation of PCR amplification of cDNA by agarose gel electrophoresis, 5 ^l of PCR product was mixed with gel loading dye (2 ml 10x TAE, 7.5 ml 40% sucrose, 830 ^l 6x bromophenol blue, 100 ml distilled water) and 3 ^l was loaded onto a 7M urea, 6% acrylamide gel (553 x 381 x 4 mm) with 1x TBE buffer and electrophoretically separated at 1500V, 50W for 2.5-3 h. The gel was transferred to filter paper, covered with plastic wrap and dried on a vacuum gel dryer (Fisher Scientific, Pttsburgh, PA) for 1 h at 80°C. After drying, bands were detected by autoradiography in a film cassette with a 33P intensifying screen. Films were exposed for 4-6 days at -80°C. All bands were manually assigned into three primary categories; constitutively expressed, differentially expressed, or fully regulated. Bands categorized as constitutively expressed were those that appeared at approximately the same density in both colonial and single cells. Bands identified as differentially expressed were those whose autoradiographic density was substantially different between colonial and single cells. Fully regulated bands were those that were either present or absent in one cell type compared to the other.
差异表达基因的鉴定后,在场的七个基因殖民细胞而不是单个细胞克隆和测序。差异表达PCR乐队被切除的干凝胶Clontech推荐的。短暂,乐队从凝胶/滤纸,放置在40 l ^ DNase - RNase-free水,覆盖着两滴无菌矿物油和加热5分钟的100°C。滤纸被丢弃。PCR引物相同的产品是re-amplified最初用来放大乐队感兴趣的。放大后,PCR扩增子进行琼脂糖凝胶上确保只有一个乐队。单一乐队被克隆到细菌质粒向量PCR克隆®®2.1使用威尼斯平底渔船TA工具包(Invi-trogen,卡尔斯巴德,CA)和测序在两个方向上使用M13启动网站如前所述(Allen et al ., 2005)。后序列组装和编辑使用向量NTI套房8软件包(InforMax Beth-esda, MD),假定的基因身份确定使用BLASTn和爆炸搜索短几乎相同的匹配(NCBI, http://www.ncbi.nih.gov)
结果
共有1365个基因片段来自单个细胞和1254个表达基因片段在殖民细胞中观察到这些研究(表2)。这些片段,表达117独特的碎片被观察到在殖民地中,并未观察到细胞的单个细胞和七个碎片殖民细胞表达但没有在单个细胞被发现。的172个差异表达基因中观察到鞭毛细胞,其中87%(149)要么是殖民细胞相比管制或打开。相比之下,只有23%(14)殖民61年观察到的差异表达基因的细胞调节或打开相对于单个细胞。
观察到的7个基因片段只在殖民细胞克隆和测序,
表3相关的差异表达基因p globosa殖民地
克隆规模(bp)基因银行最近的爆炸冲击的身份加入。基因库
加入价值预测函数。
PgC-3 490个基点DQ886384 AAP94719
PgC-10 342个基点DQ886385 ZP00777402
PgC-19 213个基点DQ886386 AY741371
PgC-13 384个基点DQ886387 XP639957
PgC-5 261个基点DQ886388 AF100332
PgC-6 486个基点DQ886389 AF107586
Emiliania huxleyi NADH 4 x 10 14
我UDP-3-0-acyl 1 x 10-37脱氢酶亚基
N-acetylglucosamine脱乙酰酶(细菌)Emiliania huxleyi CCMP的菌株373核糖体rna基因假想蛋白2 x 10 - 6
DDB01862282 (Dictyostelium discoideum)石斛兰grex夫人6 x的纯Thong-IN公认的21 d7蛋白质(ovg29)石斛兰grex夫人4 4 x打败Thong-IN假定的细胞控制蛋白(otg4)
电子传递脂质组装
蛋白质合成9 x 10 - 50
类似于蛋白质Srg6假定的应力响应
差异表达基因在兰花花卉过渡
差异表达基因在兰花植物过渡基因参与细胞分裂虽然没有产生可用的序列信息(表3)。这些片段的大小范围从213到490个基点平均362±114个基点。公认的基因识别,一个是参与能源生产(PgC-3)最近相似烟酰胺腺嘌呤二核苷酸(NADH)脱氢酶亚基即PgC-10最密切相关细菌UDP-3-0-acyl N-acetylglucosamine脱乙酰酶编码基因参与脂质大会,和PgC-19最类似于大亚基核糖体基因Emiliania huxleyi。有趣的是,剩下的三个片段似乎与应激相关蛋白或基因在高等植物开花。PgC-13最类似于一个假设的变形虫盘基网柄菌discoid-eum应激反应的蛋白质。PgC-5和PgC-6似乎与兰花开花相关基因的物种。这些基因片段的序列已经存入与加入数字DQ886384-9基因库。
确定人生阶段易受病毒感染的p . pouchetii(维尔)病毒和病毒感染和裂解的过程特征为p . pouchetii和p . globosa Bratbak et al . 1998 a, b;Brussaard et al . 2004, 2005;雅各布森2000,2002;雅各布森et al . 1996;
Veldhuis et al . 2005年)。确定哪些阶段的生命周期很容易受到病毒感染和溶菌作用是至关重要的对于理解病毒的影响棕囊藻属生命周期但是此属动态和生态。大多数以前的工作与p . pouchetii (Bratbak et al . 1998 b;哈姆et al . 1999;雅各布森2000;雅各布森et al . 1996年)和p . globosa Brussaard et al . 2004;Brussaard et al . 2005年)表明,孤独的细胞很容易受到病毒感染和细胞裂解而殖民地。然而,在一组实验中,Baudoux Brussaard殖民地(2005)表明,p . globosa细胞可以通过给出病毒感染。确定哪些生活阶段的p . pouchetii很容易受到病毒感染和细胞溶解,雅各布森et al . (2005) 2003 mes-ocosm进行了一系列的实验研究。在一组实验中p pouchetii单独细胞孵化与病毒不同数量的增加和控制不含病毒(雅各布森et al . 2005年)。 In a second set of experiments, colony cells that were detached from their colony matrix were incubated with different amounts of virus additions and a control containing no viruses; for comparison solitary flagellate cells were also included in this experiment. Colonies were shaken strongly in order to detach the cells from the mucus; this method is effective in producing colonial cells free of their mucus matrix because, in contrast to
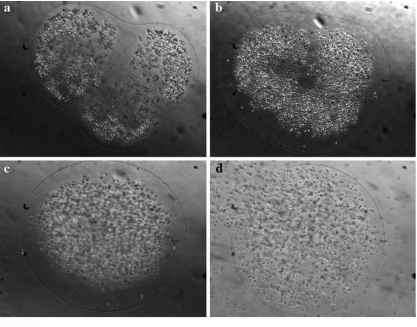
图3的时间序列中的单个p . pouchetii殖民地独立和维护好板从2003年昌盛(见Mesocosm实验方法);数据格式:最长线性尺寸和照片的日期和时间(一个)880毫米;2003年3月26日,十一33 (b) 920毫米;2003年3月27日11:57 (c) 930毫米;2003年3月28日09:41 (d) 890毫米;2003年3月29日09:19;细胞培养板在殖民地的皮肤和观察到的是能动的
p . globosa殖民地p . pouchetii容易断裂(雅各布森et al . 2005年)。
完整的殖民地孵化与病毒感染的病毒显示没有证据或裂解(雅各布森et al . 2005年)。在殖民地的实验细胞脱离他们的殖民地,独立殖民地细胞显示,没有证据表明细胞溶菌作用,而孤独的鞭毛细胞被完全溶解在实验期间(雅各布森et al . 2005年)。
在p . pouchetii殖民地形状变化伴随殖民地衰老(CSWP;RCWP)衰老的棕囊藻属殖民地已经观察到的自然现象,但是此属mesocosm和实验室条件下(戴维森和马尔尚1992;兰斯洛特和卢梭1994;卢梭et al . 1994;Veldhuis et al . 1986;真实性et al . 1988年)。禁闭室内释放从开花衰老殖民地已终止记录(Kornmann 1955;卢梭et al . 1994;真实性et al . 1988年)。然而,几乎没有详细了解一般colony-senescence和bloom-termination流程。 In an in vitro study of P. pouchetii, colonies were isolated in well plates from the 2003 mesocosms, observed over a three-day period, and digital micrographs were taken to record colony morphology and measurement of dimensions.
在这项研究中,一个单一的殖民地被拍到随着时间的推移,显示了一个典型的形态学变化小叶p pouchetii形状为球形模拟的(图3)。观察到的形状变化所需的时间间隔是两天。大约一天后观察到的形状变化,观察菌落包含能动的细胞和也发布了一些细胞中(图3 d)。而球形殖民地p . globosa的可能性存在,我们没有发现证据的存在p globosa在这些研究中(见部分在这个手稿《物种识别在Meso-cosms)。需要进一步调查文件在其他情况下观察到的衰老现象,并严格描述和量化观察形态学变化。
量化或调查的过程控制参数
率量化过程
部门(CDWP)和增长(CGWP) p pouchetii殖民地惠普尔et al。(2005 b)报道,体外平均殖民地部门mesocosm-derived率p pouchetii殖民地范围从0.21 - -0.28普通人。他们还发现,这些殖民地需要平均
3.5 - -4.9天完成一个部门;中描述从Cariou文化数据(卢梭et al ., 1994)表明,p . globosa殖民地部门发生4 - 5天后,表明相似p pouchetii(惠普尔et al . 2005 b)。惠普尔et al。(2005 b)报道,平均增长率的殖民地,作为最大线性尺寸,测量范围从0到7 | m h附近。
控制参数
硅藻细胞膜对p . pouchetii集落形成率(经验)从孤独的过渡细胞殖民形式是一个关键事件棕囊藻属生命周期。但是此属许多花朵由殖民地生活的主要阶段(但见Wassmann et al . 2005年)。甚至在近一个世纪的研究,没有明确的条件建立了单独的细胞集落形成的必要条件。硅藻细胞膜被视为集落形成粒子核,包括一系列的观察小棕囊藻属殖民地但是此属硅藻的附着在刚毛细胞膜(尤其是Chaetoceros) (Boalch 1987;兰斯洛特和卢梭1994;卢梭et al . 1994年)。卢梭et al。(1994)指出,“…实验工作在实验室控制条件下(卢梭和戴维斯,未发表的数据),给出了强有力的证据,任何微观粒子,生物(如硅藻)、有机或矿物(沙子、玻璃棉)可能作为殖民地发展”的衬底(卢梭等。1994年,35页)。Peperzak et al。(2000 b)建议,除了提供一种衬底抵消当地湍流,硅藻可能影响分泌维生素B1、集落形成的一个可能的增长刺激器,或耗尽维生素B12,假定生长抑制剂。这些观察导致假设硅藻细胞膜可能会增强集落形成率。
在2002年mesocosm研究中,进行了一个实验来确定体外p pouchetii集落形成率将由硅藻的存在增强细胞膜加入培养基;详细的方法和结果出现在Nejstgaard et al。(新闻)。集落形成率估计使用水从nutrient-amended mesocosm过滤去除棕囊藻属殖民地但是此属但允许单独细胞通过。硅藻土添加到殖民地在治疗好盘子,而控制孔板只有过滤mesocosm水(Nejstgaard et al . 2006年)。在所有的实验中,随着时间的推移新的殖民地近似线性增加。集落形成率过滤mesocosm水范围从1.3 - -1.9殖民地ml-1 d 1;利率对水dia-tomaceous地球范围从2.6 - -2.8殖民地ml-1 d 1。线性回归的斜坡与硅藻土过滤水和水是明显不同(Nejstgaard et al . 2006年)。的一般结论Nejstgaard et al .(2006)是硅藻土强化p . pouchetii体外集落形成率。
信息物质对集落形成的影响由于mesozooplankton和microzooplankton (PRI)。长和他的同事们进行了一系列的实验来测试p . globosa孤独的细胞的能力应对grazer-associated信号通过改变集落形成或蚁群的规模。集落形成评估当p . globosa遇到microzooplankton或直接mesozooplankton食草动物,只有这些食草动物的化学信号,或不吃草的线索(长2004;长等人提交)。与混合浮游动物在最初的实验中,三个主要食草动物成年桡足类的分类群Acartia tonsa和Pseudodiaptomus pelagicus(大小ca。1毫米)和异养鞭毛虫夜光虫scintil-lans;p . globosa殖民地的尺寸是直径约50 |米。在其他的实验中,a tonsa游仆虫属和种虫害是单独使用作为单独的食草动物治疗。
两种类型的容器,包括连接车厢分离膜和允许连接车厢之间的化学交换而防止藻类和食草动物在隔间之间移动。p . globosa细胞被添加到两个隔间而微或mesozoo-plankton食草动物被添加到只有一个隔间食草动物的治疗方法。一组容器是1 L塑料瓶删除和覆盖着Nitex网®使用强力胶(1 | m)。这些网格瓶放置在2 l玻璃烧杯,允许化学交换在260 cm2网。第二组的容器,称为“通信室”,是由两个0.64厘米厚,清晰、丙烯酸、封顶管(每个直径8.89厘米,长度大约为7.94厘米,每500毫升)的最后一卷螺纹到彼此,也是由一个0.8 |米分开聚碳酸酯膜滤器(长2004;长et al .,提交)。
棕囊藻属globosa细胞被添加到两个隔间,但是此属食草动物被添加到只有一个食草动物治疗。这使得同时评估食草动物的直接和间接影响的群体形成相比grazer-free控件(长2004;长et al .,提交)。加强实验之间的化学信号交换室,网状瓶子被放在板凳上高层轨道瓶和旋转50转;每个瓶子也轻轻地抬起一天两次,直到一半的体积是流离失所的从一个房间到另一个(长2004;长et al .,提交)。额外的细节的设置实验,抽样方法和结果可以在很久很久et al。(2004)和(提交)。
在最初的混合浮游动物实验中,长et al。(提交)发现甲壳纲动物mesozooplankton,由成人桡足类a tonsa和p . pelagicus显著抑制集落形成。在其他的实验中,长et al。(提交)发现信息物质线索仅从mesozooplankton a tonsa独自镇压殖民地的形成。长et al。(提交)也发现a的殖民地抑制效果tonsa密度依赖。在实验microzooplankton游仆虫属spp。长et al。(提交)报道,info-chemical线索从这个纤毛虫增强p globosa集落形成。
讨论
孤独的细胞转换殖民地
硅藻细胞膜提高p pouchetii体外集落形成(Nejstgaard et al . 2006年),这一过程可能会因此加速原位孤独的能动的细胞转变成新的殖民地。这是关键的步骤,它的形成colony-based棕囊藻属水华,但是此属,因此是至关重要的建设量化生态系统模型的棕囊藻属。但是此属未来的研究必须精心的细节由硅藻细胞膜集落形成率的控制,如剂量反应,物种差异(棕囊藻属,但是此属硅藻),以及这如何控制可能会改变在开花。一个方法,这些结果可以用机械模型包括一个集落形成率算法,集落形成率变化对棕囊藻属环境但是此属硅藻细胞膜的密度。
化学信号实验的结果指向一个潜在消费者生物体的重要作用影响棕囊藻属生命周期。但是此属唐(2003)发现microzooplankton放牧导致增加细胞的比例在殖民地生活阶段,和增加了殖民地的平均大小;长et al。(提交)还发现,信息物质从纤毛虫(Eupl-otes spp)增强在p . globosa集落形成。相比之下,长et al。(提交)发现的化学信号mesozooplankton Acartia tonsa抑制p . globosa集落形成。因为不同的反应观察中央与microzooplankton食草动物,长et al。(提交)假定p globosa可以检测放牧,识别食草动物类型,并对增强或抑制集落形成或发展现有的殖民地。这种类型的诱导反应棕囊藻属意味着一个复杂的群体形成控制机制,但是此属化学信号接收,处理,和响应依赖于信号中包含的信息。进一步的实证研究可能包括识别信号的化学物质,和决心的食草动物类群特异性信号。唐朝(2003)和长et al。(提交)结果将允许控制结构被添加到机械模型涉及到食草动物的检测,确定发送方和适当的回应的棕囊藻属单独细胞或殖民地但是此属食草动物。此外,长期的结果et al .(提交),a . tonsa的殖民地抑制效应是密度制约的,给生产一个剂量反应的第一个数据算法的影响这些grazer-derived info-chemicals包含棕囊藻属生命周期阶段但是此属浮游生物模型。这类控制演示有机体的biocom-plexity符号学的一个方面可以在生态系统模型生成。
实验结果使用粗糙地面的材料收集的浅海湾沉积物表明,底栖生物粒子可能函数作为群体形成的网站。殖民地出现在孵化期间上覆水来自细胞或非常小的殖民地与沉积物。推断,这些细胞或小殖民地自然与这些沉积物有关,但是不能排除这个可能性,他们artiW-cially混合沉积物在时间的集合。这些研究并不旨在证明或驳斥发生或存在的一个真正的底栖生物的生命周期阶段,尚未发现自原假设的存在(Kornmann 1955;凯塞1970)。相反,最简洁的解释是,这些粗糙的沉积物是附件网站发起的新殖民地,很像硅藻和硅藻细胞膜水柱(卢梭et al . 1994;雅各布森2002;Nejstgaard et al . 2006年)。殖民地整个mesocosm研究期间出现在峡湾(Nejstgaard et al . 2006年)。这些结果表明,浅层沉积物基本照明水域棕囊藻属殖民地但是此属期间增长可能作为集落形成网站。 The ecological contribution of colonies derived from such a mechanism must await a thorough seasonal study that includes sampling before and after the period of colony appearance in the water column. This Wnding suggests that some form of benthic involvement should be included in Phaeocystis life cycle models; these experiments do not provide evidence for a distinct benthic life cycle stage of P. pouch-etii. Mechanisms that provide for benthic sediments providing colony formation sites could be provisionally included in models of Phaeocystis life cycles. Empirical studies to provide details of these sediment mechanisms are required to further elaborate conceptual or mechanistic models.
比较基因表达之间徘徊单一和殖民棕囊藻属细胞表明一个更大的但是此属一系列基因表达在单级生命周期阶段比在殖民地的生命周期阶段,这意味着增加生化复杂性与单个细胞有关。虽然差异显示分析进行这些研究的一部分,绝不代表全面表征分子差异殖民和单细胞生命周期,有趣的是几个基因的表达在殖民棕囊藻属细胞,但是此属不是鞭毛单个细胞似乎与压力有关,细胞信号和复杂的发育转变高等植物。此外,基因的存在棕囊藻属最密切相关,但是此属系统不同的微生物包括细菌、变形虫、高等植物,除了密切相关haptophyte Emiliania huxleyi,表明棕囊藻属可能但是此属的基因组的基因组成的一个复杂的马赛克和监管元素类似于其他微藻,包括硅藻,他最近完全基因组测序(Montsant et al . 2005年)。
这里描述的研究也证明棕囊藻属sp.但是此属的温顺基因组和转录组研究在未来。用相对较少的修改可以适应商用RNA净化包工作与植物细胞RNA适合构建互补脱氧核糖核酸数据库从单个细胞和棕囊藻属globosa但是此属殖民地。一些先前的报道表明,基因组和质体DNA适用于分子的研究也相对容易获得各种棕囊藻属物种,但是此属(Zingone et al . 1999;兰格et al . 2002年)。发生在生物学的各个领域,从单个生物分子信息和社区的生物产生巨大的洞察生理、生活史和生态的生物包括mic-roalgae分子信息(艾伦2005年,沃克et al . 2005年)。毫无疑问,重要的新见解的生活史棕囊藻属时将获得完整的基因组但是此属和全面的基因表达数据变得可用。
殖民地扩散
体外殖民地部门文档提供了两种类型的证据在p . pouchetii殖民地扩散。首先,殖民地从孤独的细胞可能形成和成长通过有丝分裂和细胞外的分泌粘液形成了很大的殖民地观察花朵。细胞数量和生物量之间的关系已经记录了p globosa(卢梭et al . 1990年)和p . pouchetii(新闻真实性等人);这些数据将增强p . pouchetii殖民地尺寸测量的有效性。第二,现有的殖民地可能分裂,分裂后的大小也会增加通过有丝分裂和细胞外的分泌粘液(惠普尔et al . 2005 b)。后者途径使得殖民地的数量增加的速度比可能如果所有殖民地产生孤独的细胞,因为女儿殖民地由殖民地部门从数以百计的细胞,并可能获得大尺寸比殖民地建立了一个孤独的细胞快得多。与殖民地分裂的可能性,殖民地从孤独的细胞构成来源构成人口的大量的多细胞殖民地可以推导出连续的增长和殖民地范围划分阶段。建模这种殖民地扩散模式可以提供洞察其可能对棕囊藻属布鲁姆但是此属动力学的影响。需要进一步研究来确定这种模式在多大程度上是自然棕囊藻属花朵但是此属中观察到。
殖民地衰老和孤独的细胞释放
球形殖民地与支持遗传数据的观察,确定他们信心高p . pouchetii表明,菌落的形状本身应该与其他方法确认明确识别棕囊藻属殖民地但是此属的物种中观察到。殖民地的体外观察形态学改变p pouchetii表明可能有许多殖民地细胞活动和生理的变化,需要执行这个在殖民地衰老形态学改变。尚不清楚详细什么改变殖民地细胞衰老期间经历;然而,这种菌落形状改变观察点可能发生重大变化的时期。殖民地细胞特征的进一步调查这个形变期间可能会进一步了解衰老过程P。
pouchetii。定性形状变化,这些结果中描述,不能有效地纳入棕囊藻属生命周期模型,但是此属然而,如果在衰老细胞生理学的细节被发现的进一步研究这个殖民地的阶段,他们可能会证明有用的模式发展。
殖民地或单独细胞损失:病毒感染和细胞溶解
如果花朵形式主要从殖民地,p . pouchetii能逃脱病毒裂解,形成殖民地。直到殖民地经历衰老和释放孤独的鞭状的细胞,这将是预测容易受到病毒、p pouchetii殖民地细胞免受病毒感染。透明exopolymer粒子(TEP)似乎绑定病毒(Brussaard et al . 2005),引入了另一个控制衰老期间可能特别重要Phaeo-cystis殖民地的花朵。
雅各布森et al。(2005)报告说,分离殖民细胞p pouchetii似乎non-infectable。这表明蚁群细胞可能或收购一些固有的抵抗病毒感染,不涉及皮肤的殖民地。因为独立的殖民地会含有TEP殖民地粘液包围,给出粒子可能被绑定到TEP一样,减少感染分离细胞的能力。这种潜在的混杂因素应该在未来解决实验。雅各布森(2002)提出,殖民地覆盖物孔隙大小太小承认病毒粒子可能是预防病毒感染机制的殖民地。结果报道在雅各布森et al。(2005)表明,另一个机制可能负责细胞溶菌作用观察p . pouch-etii缺乏殖民地。这些实验并不是旨在阐明特定病毒感染机制,需要进一步研究来证实这些裂解的初步观察和调查机制抵抗(雅各布森et al ., 2005)。Ruardij et al。(2005)合并病毒裂解成p . globosa仿真模型;如果雅各布森et al .(2005)的结果纳入一个仿真模型的p . pouchetii病毒裂解只会涉及单独细胞一旦被殖民的形式就会刀枪不入的病毒。
摘要和结论
几个隔间,流程,和他们的控制,包括在惠普尔et al。(2005)我们理解差距进行实证分析。概念模型的两个主要方面。
第一个涉及测试生命周期过程或生产存在的这样一个过程的描述。这里讨论五个例子包括(a)的体外殖民地部门p pouchetii (CDWP);(b)的描述体外p . pouchetii与衰老有关的殖民地形状变化(CSWP);(c)决定哪些生活阶段p . pouchetii单独细胞或殖民地,很容易受到病毒感染和裂解(维尔);(d)的潜在水底的参与p pouchetii生命周期(CFBI);(e)示范单独与殖民细胞基因差异表达的p . globosa (DGE)。
第二方面涉及量化过程或监管控制。p . pouchetii率量化过程的两个例子是:(a)体外集落分裂率(CDWP)和(b)体外集落增长率(CGWP)。还有两个例子调查过程控制。首先是硅藻细胞膜增强从孤独的细胞集落形成p pouchetii(经验),第二个是抑制p . globosa的集落形成infochemi-cals mesozooplanktonic发布的食草动物,和增强的集落形成infochemi-cals microzooplankton发布的食草动物(PRI)。
调查这些特性的意义的上下文中的棕囊藻属生态学但是此属Phaeo-cystis生命周期模型是新见解和理解非常适合于在一个预先存在的组织方案,建立了事实和假设。这个新的理解,连贯的生命周期模型,可以提高生态系统模拟模型(例如,Canziani andHallam 1996;维里蒂2000;兰斯洛特et al . 2005;Ruardij et al . 2005年)旨在获得生命周期信息,涉及到更广泛的生态系统。从这个角度看,棕囊藻属是一个明确的例子,但是此属复杂自适应系统(CAS)(沃尔德罗普1992;考夫曼1993、1995)包含在其更广泛的生态系统,也被视为一个复杂的自适应系统。使用模型、概念或生态系统,推动实证研究,然后最终纳入新模型结构可以提供更好的洞察的直接和间接影响棕囊藻属数量等重要的当代问题但是此属全球生物地球化学循环,以及阐明棕囊藻属的有组织的生态系统。但是此属的角色
确认作者要感谢t . Sorlie a . Aadnesen和h Gjertsen卑尔根大学的海洋生物站在mesocosm实验寻求帮助。我们感谢现任和前任Skidaway海洋研究所的工作人员:m . Zirbel t·沃尔特斯j·丹弗斯k . Kohlberg g·斯莫利·d·希顿,j . Brofft和r . Hristov。我们还要感谢现任和前任乔治亚理工学院的学生和教职工:t Barsby和m .干草。资金是由美国国家科学基金会提供0 - 00 - 83381页。本文对再保险公司工作组的贡献120“棕囊藻属,但是此属主要环节climate-relevant元素的生物化学循环”。raybet雷竞技最新
引用
艾伦AE (2005) DeWning能量平衡的分子基础海洋硅藻Xuctuating环境条件下。J Phycol 41:1073 Allen AE - 1076展位MG,真实性PG,弗里希我(2005)InXuence硝酸可用性的分布和丰富的异养细菌硝酸盐同化基因在巴伦支海的夏天。Aquat活细胞生态39:247 - 255 Ayers GP,吉列RW (2000) DMS在遥远的海洋大气及其氧化产品。J海Res 43:275 - 286
Baudoux得了,Brussaard CPD(2005)描述diVerent病毒感染海洋有害藻华棕囊藻属globosa但是此属的物种。病毒学341:80 - 90 Boalch GT(1987)最近的花朵在西方英语频道。拉普pv Reun缺点int空洞Mer 187:9497
Bratbak G,雅各布森,Heldal M(1998)病毒裂解的棕囊藻属pouchetii但是此属和细菌二次生产。Aquat活细胞生态16:11-16 Bratbak G,雅各布森,Heldal M,长崎K, Thingstad F (1998 b)病毒生产棕囊藻属pouchetii但是此属及其与宿主细胞生长和营养的关系。Aquat活细胞生态16:1-9 Brussaard CPD, Kuipers B, Veldhuis MJW (2005) mes-ocosm棕囊藻属globosa但是此属种群动态的研究即监管作用的病毒盛开控制。有害藻类4:859 - 874 Brussaard CPD,短的SM,完了厘米,净重的CA(2004)的隔离和系统发育分析小说《浮游植物病毒感染棕囊藻属globosa但是此属(Prymnesiophyseae)。:环境Microbiol 70:3700 - 3705
Canziani GA,哈勒姆TG(1996)数学模型对棕囊藻属sp.但是此属浮游生物群落动态控制。我基本模型非线性世界三19 - 76
Cariou V, Casotti R, Birrien杰,Vaulot D(1994)的起始棕囊藻属殖民地。但是此属J浮游生物Res 16:458470
戴维森,Marchant HJ(1992)棕囊藻属(Prymnesiophyceae)。但是此属的生物学和生态学:铁,查普曼DJ (eds) Phycological研究的进展。Biopress有限公司,英国布里斯托尔、pp 1-45 Eilertsen HC(1989)棕囊藻属pouchetii但是此属(Hariot) Lagerheim北极海洋生态系统的一个关键物种:生活历史和生理学。拉普pv Reun缺点int Ex-plor Mer 188:131弗里希我,汉森,威利是的,Wimbush J,穆雷J, Nierzwicki-Bauer SA (2002) 18 s rRNA基因的特定放大方法检测斑马贻贝(Dreissena polymorpha)幼虫在浮游生物样品。Hydrobiologia 487:33-44 Gabric AJ, Matrai PA,韦尔M(1999)模型的生产甲基在巴伦支海的春天的花朵。忒勒斯爵士B化学phy Meteorol 51:919 - 937
Guillard RRL,哈格雷夫斯体育(1993)Stichochrysis immobi-lis硅藻,不是金藻。Phycologia 32:234236
哈姆CE, Simson哒,默克尔R,斯V(1999)殖民地的棕囊藻属globosa但是此属受薄但艰难的皮肤保护。3月生态掠夺Ser 187:101 - 111雅各布雷竞技csgo森(2000)新方面的开花动态棕囊藻属pouchetii但是此属(Haptophyta)在挪威水域。博士学位论文。渔业和海洋生物学(45页)。挪威卑尔根大学卑尔根,雅各布森(2002)形态,相对DNA含量和假想的生命周期的棕囊藻属pouchetii但是此属(Prym-nesiophyceae);特别强调鞭毛细胞类型。Sarsia 87:338 - 349雅各布森,Bratbak G, Heldal M(1996)病毒感染的隔离和表征棕囊藻属pouchetii但是此属(Prymnesiphyceae)。J Phycol 32:923 - 927雅各布森,Martinez-Martinez J,真实性P,弗里希我,Sandaa RA,拉森(2005)殖民地或殖民细胞Phaoecystis pouchetii (Prymnesiophyceae) su-septible病毒感染吗?美国社会的湖沼学和海洋学,夏季会议。(西班牙圣地亚哥,考夫曼SA(1993)秩序的起源。 Self-organization and selection in evolution. Oxford University Press, New York Kauffman SA (1995) At home in the Universe. The search for laws of self-organization and complexity. Oxford University Press, New York
凯塞H(1970)研究调查在棕囊藻属poucheti但是此属(Haptophyceae)。Helgol威斯康星州Meeresunters 20:195 - 212 Kornmann PV (1955) Beobachtungen棕囊藻属,但是此属
Kulturen。兰斯洛特Helgol威斯康星州Meeresunters 5:218 - 233 C,凯勒博士卢梭V,史密斯我们Mathot年代(1998)海洋Haptophyte棕囊藻属sp。但是此属个体生态学:安德森DM, Cembella广告,Hallegraeff通用(eds)有害藻华的生理生态学。兰斯洛特柏林斯普林格出版社,页209 - 224 C,卢梭V(1994)生态学的棕囊藻属,但是此属殖民地形式的关键作用。:绿色JC,里德比特BSC (eds) haptophyte藻类。克拉伦登出版社,牛津兰斯洛特pp 229 - 245 C,施皮茨Y, Gypens N, K•鲁迪,Becquevort年代,卢梭V, Lacroix G, Billen G(2005)模拟硅藻和棕囊藻属开花,但是此属的养分循环的南湾北海:米罗模型。3月生态掠夺Ser 289:63兰斯洛特- 78 C, Wassmann P,巴斯H(1994)生态学的阶段
ocystis-dominated生态系统。J 3月系统5:1-4兰格M,陈Y-Q Medlin路(2002)分子遗传界定棕囊藻属物种,但是此属(Prymnesiophy-ceae)使用编码和非编码区域的核和质体基因组。欧元J Phycol兰格37:77 - 92 M, Laure G Vaulot D,西蒙N,阿曼RI,路德维希W(1996)识别类Prymnesiophyceae和属棕囊藻属,但是此属核糖体RNA-targeted核酸探针检测到流式细胞术。J Phycol 32:868 - 872长JD(2004)的可塑性consumer-prey交互在海里:化学信号,厌恶学习,和生态后果。博士学位论文。生物学系(119页)。乔治亚理工学院,亚特兰大,乔治亚州,美国。URL链接:http://www.etd.ga tech.edu/theses/available/etd - 11雷竞技csgo182004 - 164652/unre stricted/long_jeremy_d_200412_phd.pdf长JD,斯莫利GW, Barsby T,安德森JT,干草我(提交)化学线索诱发特定于使用者的防御在b
这篇文章有用吗?