棕囊藻属电子但是此属
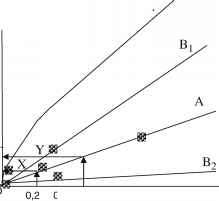
图4的计算分子钟。(a)所示的ml树Edvardsen et al。(2000)线性化(Kooistra和Medlin 1996),这样所有的进化率是相同的。化石可以追溯到coccolithophore类群被放置在这些类群有其Wrst节点出现在化石记录(打开树圈)。(b)分支长度对化石的回归日期已经完成根据Kooistra Medlin (1996)。从这个回归线我们推断的散度温暖和冷水棕囊藻属(Y),但是此属的散度p pouchetii从南极洲p (x)(固体圈树)。(c)白垩纪以来南极表层海水温度与分子树绘制成比例的时间和温度,重新从起重机械(1993)
从冷水温水棕囊藻属的物种分化但是此属物种大约30 Ma,伴随着时间的德雷克海峡打开和ACC系统成立。这将有效地孤立的祖先种群在南极足以让他们从他们的温水形成物种的祖先。南极洲的分离从p p . pouchetii大约是15,恰逢一个重大气候变暖事件在世界上的海洋(图4)。在此之前的数量必须能够穿过赤道从南到北,因为水温冷却可以生存,但这变暖事件分离两个极地的人群让他们发散到我们今天的两个物种在两极。发现了类似的结果为foraminfera(亲爱的et al。2000年,2004年)。
因此分子数据定义了我们物种和建议哪些物种很可能是由神秘的物种。下面我们详细的基本描述每个物种和描述的正式提供一些指示其他未定种,这个信息是已知的。
正式描述物种
发布的有效分类单元的概述近年来一直在审查提出了以下。这包括物种属在最近的评论中列出(Sour-nia 1988)和新物种描述日期之后,除了三个类群,并没有被研究过,因为他们的描述,p . amoe-boidea Buttner, p . sphaeroidea Buttner和p . brucei曼京。这些没有的前两个属的特征特点,Haptophyta分工,所以很可能只有后者棕囊藻属。但是此属的物种可能仍然是一个有效的物种的主要特色物种包括在本节表总结了雅各布森(2002)。
p . pouchetii (Pouchet Hariot) Lagerheim (Pouchet 1892)形式的云雾状殖民地与细胞包4(图5)。分子数据表明,这是一种复杂的但这里很少菌株与分子技术研究了所以这只是一个非常初步的建议。
鞭毛的p . pouchetii阶段(图6)是最近的形态学研究的主题(雅各布森2000,2002)。细胞圆形,平均直径5 im。鞭毛是相等的长度,ca 11 | m, heterodynamic。的haptonema非常短,1 - 2 |米,有轻微肿胀,并与光学显微镜不容易看到。身体尺度有两种类型:近圆形平面板,0.24 x 0.25 |米,和提出的边缘,形成外部层和较小的椭圆板和稍弯曲的边缘,0.19 x 0.15 | m,在更大的尺度。两种类型的刻度显示薄辐射山脊。丝(30 |米)安排在五组与典型的五角近端结构被认为在细胞外,或盘绕在囊泡在细胞表面。硅的存在是报道这些细丝(雅各布森2002)。的超微结构类似于p globosa(帕克et al . 1971年)和其他的棕囊藻属,但是此属这个信息是可用的(Zingone et al . 1999),细胞核位于后方,两个叶绿体嵌入式中,它们之间的高尔基体。这个细胞阶段可以被病毒感染(雅各布森et al . 1996年)。
p . globosa Scherffel (Scherffel 1900)形成球状殖民地与细胞均匀分布在整个殖民地(图5 b)。分子数据和DNA内容表明,这是一个复杂的三个或四个神秘的物种,但迄今为止不存在形态调查来支持这个。
鞭毛的p . globosa阶段(图6 b, c)首次描述了帕克et al .(1971)的名义p . pouchetii时这两个物种被认为是在同一物种的生命周期阶段。细胞3 - 6 |米,3和4.5 |米之间更频繁。两个Xagella和haptonema摆脱萧条细胞体内。鞭毛是相等的长度,1.5 x细胞长度和heterodynamic。haptonema是1/4到1/3 Xagella的长度。stiV和有一个明确的远端肿胀。在活细胞haptonema很容易看到,它是直接向前而细胞移动。身体尺度显示所有辐射脊表面和有两种类型:近圆形死者纪念雕柱板,0.18 x 0.19 |米,和提出的边缘,形成
-
 Phaeocystis sp.2 (a, b) taken from http://www.jo-Phaeocystis. (a) P. pouchetii, (b) P. globosa, (c) P. antarc- chemnet.de/fiu/OCB3043_21.html Scale bar = 100 ^m tica, young colony, (d) P. antarctica, older colony, (e) P.")
- 图5光jahnii的殖民地的微观显微图阶段,(f)棕囊藻属sp.2但是此属(a, b)来自http://www.jo-Phaeocystis。(a) P . pouchetii (b) P . globosa (c) P antarc - chemnet.de /事业/ OCB3043_21.html酒吧= 100 m ^行业规模,年轻的群体,(d) P南极洲,老殖民地,P (e)。
 Phaeocystis sp.2 (a, b) taken from http://www.jo-Phaeocystis. (a) P. pouchetii, (b) P. globosa, (c) P. antarc- chemnet.de/fiu/OCB3043_21.html Scale bar = 100 ^m tica, young colony, (d) P. antarctica, older colony, (e) P.")
外部层和较小的椭圆盘,0.10 x 0.13 | m和强烈弯曲的边缘,在更大的尺度。超微结构是典型的属,有两个金褐色的叶绿体,与内部梭状中,一些折射的囊泡可能包括存储产品。核后,而高尔基体,与大量的堆叠cysternae在叶绿体和核之间的空间。鞭毛基地有典型的远端和近端盘子的过渡区。的两个克隆观察到帕克et al .(1971)(147年克隆)形式长的细丝(20 |米)在五组出院外,与近端结束安排在一个非常典型的五角结构,包围着一个微弱的泡孔的中心。细胞的未偿清的线程被发现在一个泡在细胞表面。Chretiennot-Dinet et al。(1997)表明,这些细丝包含alpha-chitin。
p .南极卡斯滕(Karsten 1905)是最不知道棕囊藻属的但是此属形态学的观点。它也形成球状殖民地在殖民地与细胞随机分布的表面(图5 c, d)。这些殖民地可以变得相当扭曲和细长的随着年龄的增长。
鞭毛阶段p .南极(图6 d)已收到很少研究。只有一个可用的天平从南极棕囊藻属,但是此属插图(拉森和Moestrup 1989),显示两个不同大小的椭圆形尺度(0.27 x 0.19 im和0.18 x 0.14 | m,分别)。最近,三个不同的变种已经从字段说明材料从南极有鳞片中diVerent大小相比,拉尔森和Moestrup haptonema没有隆起的提示(Marchant et al . 2005年,无花果。6 f, 7 d-f)。尺度还发现不同大小的鞭毛阶段从隔离的殖民地,获得phylogeneti-cally接近SK 22 (Zingone和蒙特莎,未发表)。分子数据发布到日期从鞭毛虫只来自殖民阶段或阶段,最初殖民地。只有一种类型的序列施压
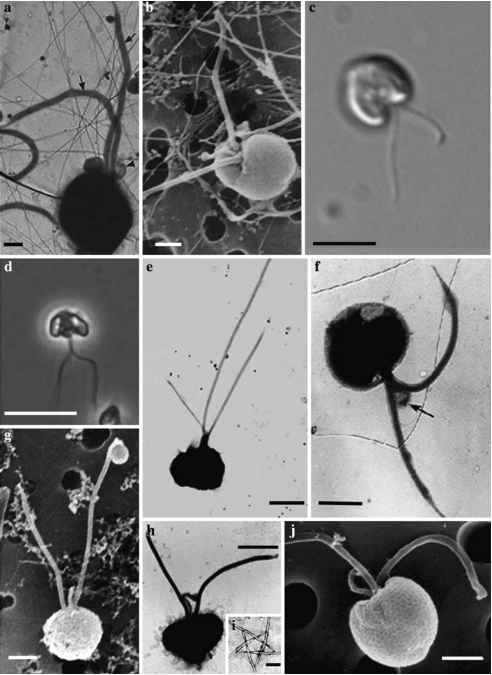
图6光学显微镜(LM)、透射电子显微镜(TEM)和扫描电镜(SEM)鞭毛的棕囊藻属。但是此属阶段的显微图(a) p pouchetii、TEM、(b) p . globosa SEM, (c) p . globosa LM, (d) p南极洲,LM, (e) p . jahnii TEM, (f)棕囊藻属sp.从南极海域TEM,但是此属(g)棕囊藻属sp。3,但是此属SEM、(h) p . cordata TEM (i) p . cordata星形模式
图6光学显微镜(LM)、透射电子显微镜(TEM)和扫描电镜(SEM)鞭毛的棕囊藻属。但是此属阶段的显微图(a) p pouchetii、TEM、(b) p . globosa SEM, (c) p . globosa LM, (d) p南极洲,LM, (e) p . jahnii TEM, (f)棕囊藻属sp.从南极海域TEM,但是此属(g)棕囊藻属sp。3,但是此属SEM、(h) p . cordata TEM (i) p . cordata星形模式five-filament的中心结构,TEM (j) p . cordata SEM。从雅各布森(a) (2002), (c)由安娜Noordeloos, (d)采取p . Assmy博士(f)的斯科特和马尔尚(2005),(e、h、i)从Zingone et al . (1999), (b)和(g) Vaulot et al。(1994)。比例尺= 1 ^ m (a, b, g)规模的酒吧= 10 ^ m (c, d)比例尺= 2 ^ m (e-j)规模的酒吧= ^ 0.3 m (i)
-
 Phaeocystis sp. 1 (PML 559), LM, (b) Phaeocystis sp. 1 (PML 559), SEM, (c) Tip of the tube-like structure ejected from Phaeocystis sp. 1 (PML 559), TEM. (d-f) Morphs 1, 2, 3 of P. antarctica, (g) filaments of P.")
- 图7 LM、TEM和SEM显微图的鞭毛的棕囊藻属。但是此属阶段(一)棕囊藻属sp。但是此属1 (PML 559), LM, (b)棕囊藻属sp。但是此属1 (PML 559),扫描电镜,(c)的管状结构逐出棕囊藻属sp。但是此属1 (PML 559), TEM。(d-f)变种1、2、3 P的南极洲,(g)丝的P。
 Phaeocystis sp. 1 (PML 559), LM, (b) Phaeocystis sp. 1 (PML 559), SEM, (c) Tip of the tube-like structure ejected from Phaeocystis sp. 1 (PML 559), TEM. (d-f) Morphs 1, 2, 3 of P. antarctica, (g) filaments of P.")
花在每个应变与多个变种的p . pouchetii和p . globosa这强烈表明,殖民棕囊藻属从南极不是但是此属物种复杂。相似scrobiculata (b, c)由Gandi馥兰妮(d-f)斯科特和马尔尚(2005)。(g)www.marbot.gu.se SSS /其他/ Phaeocystis_scrobiculata.GIF。比例尺= 10点(a)规模的酒吧= 2点(b), (g)比例尺= 0.5点(氟)
在其序列从南极菌株表明他们可能是一个单一的物种,而这些新单细胞变种在现场样本可能属于物种还不文明的
(图7 d-f)。另外,不同的鞭打形态类型可能属于不同的sub-clades antarctica-clade页。所有这些信息表明形态多样性和高表明可能有一个以上的物种在棕囊藻属从南极但是此属。平行形态调查上的菌株属于不同的p .南极洲sub-clades。
p . jahnii Zingone(图5 e)形式不同于其他棕囊藻属殖民地但是此属殖民地(Zingone et al . 1999年)。这些都是松散的聚合non-motile细胞嵌入在粘稠的粘液多糖性质的矩阵可能,没有外部层也没有一个明确的形状。在殖民地文化材料可能和利润形成宽表有时坚持细胞管。殖民细胞从6到8.5 |米24叶绿体。
鞭毛细胞p jahnii(图6 e)是圆形的,直径3.5 - 5 im,鞭毛的明显不平等的长度(-12 m和5.5 | -6.5 | 8.5 m,分别)。haptonema相对较长(3 - 4.5 im)和无明显隆起。相比其他棕囊藻属的物种,但是此属尺度更薄、更精致,与一个非常微弱的辐射模式缺乏规模的核心部分。大尺度(0.35 x 0.28 im)没有举起的边缘,而背后的小尺度(0.18 x 0.14 |米)有典型的弯曲边缘。折射的橘黄的身体通常被认为在活细胞中叶绿体之间的空间。丝没有观察到在这个物种。
p . cordata Zingone et Chretiennot-Dinet(图6 h-j)只有单个细胞的发生通常是三角形的,心形的或椭圆形,稍扁平,深鞭毛抑郁和或多或少地指出antapical结束(Zingone et al . 1999年)。平均尺寸是3 - 3.5我长,3 - 4 |米宽,2.5我厚。两个鞭毛略几乎相等的,5.5 - -7.5和4.5 6 |米长度,分别。haptonema很短(2.2 | -2.5米)在光学显微镜和几乎不可见,与膨胀结束在电子显微镜下观察到。细胞通常游泳与鞭毛杆向后,连续两个鞭毛,完全隐藏haptonema。绕纵轴的细胞,而移动。很少的细胞被认为与鞭毛杆向前移动。较大和较小的尺度都是椭圆形,m和0.18 x 0.13 x 0.18 | 0.25 | m,分别。更大的尺度上有提高的钢圈和轻微的中央旋钮,并形成细胞外部投资。小尺度内折的钢圈,形成一个内部层相邻的质膜。 The filaments are seen in disk-like vesicles underneath the cell surface (up to three in a cell) or discharged, with the typical five-ray star pattern (Fig6i). Ultrathin sections show the two flagella and the haptonema inserted along a line that is transversal to the plane crossing the plast-ids. Comparable information is not available for other species yet. The internal microanatomy is similar to that of the other species of the genus.
p . scrobiculata Moestrup(图7 g)描述了从现场收集到的材料在新西兰海域unicell (Moestrup 1979)。没有证据表明它使殖民地也没有任何分子的工作已经完成。与两种类型的细胞直径8 im的尺度,m和0.19 x 0.21 x 0.45 | 0.6 | m大小。这两种类型的尺度是无定形的背,但是随着脊辐射从平原中心腹一侧。其鞭毛和haptonema长度的两倍中发现p . globosa和尺度大约两倍。另一个特色是它产生的细丝,在九组(8对和一个),与五单纤维的生产从其他物种产生细丝(图7 g)。丝的中心模式相当不规则,不形成恒星的形状特征在中间。丝安排相同的模式也被发现在澳大利亚水域(Hallegraeff 1983)和在地中海(Zingone et al . 1999年)。然而,规模较小的澳大利亚和地中海标本,这表明可能更高的多样性是探索在这个分类单元。
未定的物种
除了在文献中描述的物种,一些类群归因于棕囊藻属,但是此属目前形态和分子调查提出了以下。
559年棕囊藻属sp。但是此属1 (PML)(图7 a - c)似乎只出现在单个细胞。两个鞭毛-12 m和5.5 | -6.5 | 8.5 m,分别。haptonema 3 - 4.5 |米长,没有凸起。更大的尺度上类似于p . cordata虽然大(0.35 x 0.22 |米),厚厚的抬起rim和中央旋钮。小尺度(0.25 x 0.17 im)向内弯的钢圈。这一物种的一个不寻常的特性是它产生的管状结构和特殊的结束被细胞。这些尸体可能存在于每个细胞的5 - 7数量(图7 b),留下一个大萧条一次挤压。多的底栖生物阶段形成年龄和文化很可能前面身体有助于把细胞基质。
棕囊藻属sp。但是此属2(图5)是迄今为止唯一Phaeocys-tis培养从地中海一直显示,形成典型的殖民地的球形(Zingone、Borra馥兰妮和Pro-caccini准备)。鞭毛虫的形状不规则,鞭毛极明显的肩膀。鞭毛是明显不平等的长度,haptonema没有膨胀的结束。没有尺度上检测到细胞表面,也没有任何类型的细丝。18 s分析证明该分类单元属于p globosa分支,虽然它由九个碱基对,潜水员的diVerence与p . pouchetii和p之间的南极洲。其序列unalignable与那些可用的其他p . globosa菌株。
棕囊藻属sp。但是此属3(图6 g)包括菌株孤立于地中海西北部。单细胞和p .规模cordata形态相似,这是初步归因于后者物种(如菌株MEDNS2和MEDNS3 Zingone et al . 1999),但形态diVerences,最初的赏识。相比p . cordata棕囊藻属sp。但是此属3略大一些,身体呈圆形,短鞭毛和更大的身体尺度几乎是圆形而不是椭圆形。初步的分子分析放置在p globosa复杂。
很明显,近年来遇到的物种形态细节以外的原始描述属的棕囊藻属,但是此属因此我们觉得有必要修订属描述如下:
棕囊藻属Lagerheim 1893,但是此属Zingone和Medlin校正。
能动的细胞有两或多或少等于鞭毛和一个短non-coiling haptonema;1 - 4顶叶叶绿体;胞体经常覆盖着平两个不同大小的尺度。Ejectile细胞器已知的几个物种。复杂的生命周期,涉及non-motile的形成阶段,不知道对所有物种。Non-motile细胞通常没有附件和尺度,单一或排列成球形,浅裂的,铠装或不规则的凝胶状的殖民地;如果附件存在,通常短或不完整。
前景
从观察到目前为止,包括字段和培养材料和分子数据,很明显,我们已经走了很长的路从10年前,当我们只有一种棕囊藻属,但是此属p globosa世界性的分布。我们现在有一个更清晰的照片属的物种及其分布。但是棕囊藻属仍然持有许多谜团。但是此属显然,有更多种类的棕囊藻属目前正式承认。但是此属这些形态是不同的,而另一些需要进一步研究来评估他们是否神秘的物种,或者更确切地说,它们形态不同的至少在某些阶段的生命周期。新途径的分子和形态调查关注分类群只知道从现场材料,如p .南极洲的三个形态类型,和还未耕作的罕见的p . scrobiculata,或从南非水域的鞭毛虫的杯状容器板(1991年皮纳尔,1996)。鞭毛虫阶段似乎更广泛和多样的殖民阶段相比,材料应该聚集通过特定的栽培技术研究(如连续稀释技术)。而薄丝的功能尚未完全阐明,独特的角色extrusomes发现559年厚度更是难以理解。或许这些diVerent附件机制,但它们也可以参与潜在过冬阶段形成(喋喋不休地说等人未发表的观察)。迄今为止使用的分子工具大大促进了描绘棕囊藻属的物种。但是此属 What remains to be clarified is the genetic diversity within the major species and how this diversity changes in time and space, which will require the set up of new high-resolution methods (see Gabbler et al. (2007)) for the latest developments in population genetic analysis of Phaeocystis). This information, coupled with a better circumscription of species, is the prerequisite for significant advancements in the understanding of the ecology of one of the key players of the world ocean's plankton.
菲利普博士确认Assmy请提供p .南极洲的照片。Gandi馥兰妮的照片提供棕囊藻属sp。但是此属1 (PML 559)。照片从图5.2斯科特和马尔尚(2005)与许可复制F.J.斯科特和H.J.马尔尚(Eds), 258年南极海洋原生生物,(2005),版权澳大利亚生物资源的研究,澳大利亚南极分裂和安德鲁·戴维森。数据从形态学、相对DNA复制内容和假设的生命周期棕囊藻属pouchetii但是此属(Prymnesi-ophyceae);特别强调Xagellated细胞类型的由Sarsia雅各布森(2002)www.tandf.no sarsia、2002、87:338 - 349年,泰勒和弗朗西斯的许可。图2 a、g Vaulot et al。(1994)和图6,9日,32 Zingone et al。(1999)与Phycological许可复制美国的社会。本文属于欧盟范围的网络卓越MARBEF(海洋生物多样性和生态系统功能)。
引用
Batje米,米歇利斯H(1986)棕囊藻属pouchetii花朵但是此属东弗里西亚沿海水域(德国湾、北海)。3月93:21-27鲍曼MEM杂志,杨克海洋Planktonalgen der Arktis J (1986)。即死Haptophycee棕囊藻属pouchetii但是此属。Mikrokosmos 75:262 - 265鲍曼MEM Brandini FP,斯桃波R(1993)的influ-cence光线和温度对碳特定DMS-release棕囊藻属南极洲和三个南极但是此属硅藻的文化。3月化学45:56 - 78鲍曼MEM,兰斯洛特C, Brandini FP, Sakshaug E,约翰DM(1994)的分类标识国际化prymnesiophyte棕囊藻属,但是此属形态和的地步的方法。J 3月系统5:23-39 Buma AGJ, Bano N, Veldhuis MJW,伊瓦(1991)比较两种色素的prymnesiophyte棕囊藻属sp。但是此属Neth J海Res 27:173 - 182
Buttner J(1911)死farbigen Flagellaten des基尔港口。威斯康星州Untersuch, NF Abt基尔12:119 - 133
常跳频(1983)粘液生产棕囊藻属pouchetii(但是此属Prymnesiophyceae)培养的1981“塔斯曼湾泥”。N Z J Mar Freshw Res 17:165 - 168
Chretiennot-Dinet MJ, Giraud-Guille mm, Vaulot D, Putaux J-L,齐藤Z, Chanzy H(1997)的几丁质的性质的细丝驱逐棕囊藻属(Prym-nesiophyceae)。但是此属J Phycol 33:666 - 672起重机械JA(1993)纬度的范围波动在海洋领域通过地质时期。趋势生态另一个星球8:162 - 166
追逐MW,克纳普年代,考克斯AV,克拉克森JJ, Butsko Y,约瑟夫·J Savolainen V, Parokonny 2003。分子系统学、吉斯和混合类群的起源在烟草(茄科)。安机器人92:107 - 127亲爱的KF,韦德厘米,斯图尔特IA,克朗D,丁格尔R,布朗AJL(2000)分子的基因混合的证据北极和南极近极的浮游生物有孔虫的数量。自然405:43-47亲爱的KF,库希拉M, Pudsey CJ,韦德厘米(2004)分子证据链接在极地浮游原生生物的多样化第四纪气候动力学。raybet雷竞技最新PNAS 101:7657 - 7662戴维森(1985)的生物学方面的棕囊藻属pouchetii但是此属(Prymnesiophyceae)(荣誉。论文)。塔斯马尼亚大学的戴维森,H -马尔尚(1992)棕囊藻属(Prymnesiophyceae)。但是此属的生物学和生态学:铁,查普曼DJ (eds) phycological研究进展,卷。8。Biopress、布里斯托尔、pp 1-45 Edvardsen B,艾克瑞姆刚W,绿色JC,安徒生RA, Moon-Van Der Staay SY, Medlin路(2000)。系统发育重建Haptophyta推断的核糖体rna序列和可用的形态学数据。 Phycologia 39:19-35 Fevolden SE, Schneppenheim R (1989) Genetic homogeneity of krill (Euphausia superbaDana)在南大洋。极地生物9:533 - 539喋喋不休地说,海耶斯PK, Medlin路(2007)方法揭示遗传多样性的集落形成prymne-siophytes棕囊藻属南极洲,但是此属p globosa和p . pouchetii——初步结果。生物地球化学(本卷)doi 10.1007 / s10533 - 007 - 9084 - 4 Hallegraeff通用汽车(1983)Scale-bearing和有甲壳的浮游生物从东澳大利亚的电流。机器人Mar 36:493 - 515
哈姆C, Simson哒,默克尔R,斯V(1999)的殖民地棕囊藻属globosa但是此属受薄但艰难的皮肤保护。3月生态掠夺Ser 187:101 - 111杨克J雷竞技csgo(1989)增长率对光线和温度的依赖关系和元素组成棕囊藻属globosa但是此属Scherffel和p . pouchetii (Har)。Lagerh。在批处理文化。Neth。J海Res 23:15-21杨克,鲍曼MEM(1986)死海洋Planktonalge棕囊藻属globosa但是此属:明信片Massenform unser Kustengewasser。杨克Mikrokosmos 75:357 - 359 J,鲍曼M(1987)区分棕囊藻属pouchetii但是此属(Har)。Lagerheim和棕囊藻属globosa但是此属Scherffel。即殖民地的形状和温度公差。Hydrobiol牛21:141 - 147
雅各布森(2000)新方面的开花动态棕囊藻属pouchetii但是此属(Haptophyta)在挪威水域。博士论文。挪威卑尔根大学,138 pp雅各布森(2002)形态,相对DNA含量和假想的生命周期的棕囊藻属pouchetii但是此属(Prymnesiophyceae);特别强调鞭毛细胞类型。Sarsia 87:338 - 349雅各布森,Bratbak G, Heldal M(1996)病毒感染的隔离和表征棕囊藻属pouchetii但是此属(Prymnesiophyceae)。J Phycol Karsten 32:923 - 927 G (1905) Das浮游植物des Antarktischen米尔新一轮材料der德国TiefseeExpedition 1898 - 1899。威斯康星州。Ergeb。多伊奇。Tief。 Exp. Valdivia 1898-1899 Band I. , Teil 2, 136 pp Keller MD, Ellows WKB, Guillard RL (1989) Dimethyl sulfide production in marine phytoplankton. In: Saltz-mann E, Cooper W (eds) Biogenic Sulfur in the Environment. American Chemical Society, Washington DC, pp 167-182 Kooistra WHCF, Medlin LK (1996) Evolution of the diatoms (Bacillariophyta): IV. A reconstruction of their age from small subunit rRNA coding regions and the fossil record. Mol Phyl Evol 6:391-407 Kornmann P (1955) Beobachtungen an Phaeocystis-
Kulturen。Helgol威斯康星州米尔Lagerheim 5:218 - 233 G(1893)棕囊藻属11月但是此属grundadt pa
四孢藻属poucheti Har。那不是1:32-33兰斯洛特C, Billen G, Sournia, Weisse T, Colijn F, Vel-dhuis MJW,戴维斯,Wassmann P (1987) Phaeo-cystis花朵和营养浓缩在北海大陆沿海地区。中16:38-46
兰格M(1997)内的分子遗传学调查属棕囊藻属(Prymnesiophyceae)。但是此属P.D.论文不莱梅大学、德国、170页兰格M,陈Y-Q Medlin路(2002)分子遗传界定棕囊藻属物种,但是此属(Prymnesiophy-ceae)使用编码和非编码区域的核和质体基因组。欧元J Phycol 37:77 Larsen J - 92 Moestrup 0(1989)指南毒性和潜在的有毒海藻。鱼检验服务,渔业部长哥本哈根Medlin路,兰格M,鲍曼MEM(1994)三个克隆形成物种之间的遗传分化棕囊藻属,但是此属Prymnesiophyta发展史的进一步证据。Phycologia 33:199 - 212 -马尔尚HJ,戴维森,凯利GY(1991)紫外线保护化合物在南极洲的海洋藻类棕囊藻属pouchetii但是此属。3月柏林109:391杂志- 395年马尔尚HJ,斯科特•FJ戴维森圣(2005)Haptophytes: Prymnesiales秩序。:斯科特•FJ -马尔尚HJ (eds)南极海洋原生生物。堪培拉,澳大利亚生物资源研究pp 255 - 275 Moestrup 0(1979)电子显微镜鉴定的海洋微型浮游生物从新西兰包括四个新物种的描述。N Z J机器人17:61 - 95
Moestrup 0,拉森J(1992)潜在的有毒浮游植物1。Haptophyceae (Prymnesiophyceae)。:年代林德利(ed)冰对浮游生物识别传单,传单
179号。自然环境研究委员会,普利茅斯,页1 - 11,蒙特莎米洛夫乔伊C,奥尔西尼L, Procaccini G(2003)双相的分布cyst-forming腰鞭毛虫Polarella glacialis。极地生物Olbers 26:186 - 194 D, Gouretski V,检波器G,葵花子粉J (1962) Hydro-graphic南大洋的阿特拉斯。Druckhaus Nord:不莱梅哈芬。82板帕克,绿色JC,曼顿我的精细结构(1971)观察zoids属的棕囊藻属(Haptophyceae)。但是此属英国51:927 - 941 J Mar Assoc皮纳尔RN(1991)线程的能动的细胞形成
棕囊藻属。但是此属电工实习。Microsc Soc年代非洲21:135 RN - 137皮纳尔(1996)观察棕囊藻属在南非水域但是此属的盘轴承阶段。在:Borg M, Semesi Pederson M,伯格曼B (eds)目前的趋势在东非地区的海洋植物研究(77 - 98)Sida,海洋科学计划,部门的研究合作,SAREC。ISBN > 91-630-4907-4 Pouchet G(1892)关于algue pelagique中篇小说。
考虑建筑渲染降神会16 Janvier 44:34-36卢梭V, Vaulot D, Casotti R, Cariou V,楞次J, Gunkel J,鲍曼M(1994)的生命周期棕囊藻属(Prymnesiophyceae):但是此属证据和假设。J 3月系统5:23-39 Scherffel(1900) 11月规格棕囊藻属globosa但是此属nebst并把Betrachtungen uber Phylogenie倪德勒死去,insbesondere布劳恩Organismsen。威斯康星州米尔Abt Helgo 4:1-28
Schoemann V, Becquevort年代,Stefels J,卢梭V,兰斯洛特C(2005)全球海洋棕囊藻属花朵但是此属及其控制机制:一个回顾。J海Res 53:43 - 66
斯科特•FJ -马尔尚HJ(2005)南极海洋原生生物。
澳大利亚生物资源研究,堪培拉沈P范Rijssel M,王Y, Songhui L, Jufang C,气Y(2004)从中国有毒棕囊藻属globosa但是此属菌株生长在非常高的温度。:Steidinger KA,兰茨贝格JH,托马斯CR、Vargo GA (eds)有害藻类2002。佛罗里达鱼类和野生动物保护委员会,佛罗里达理工学院的海洋学和联合国教科文组织的政府间海洋学委员会,圣彼得堡,页396 - 398史密斯我们科迪斯波蒂洛杉矶,纳尔逊DM,引发T, Buskey EJ, Niebauer HJ, GF(1991)科棕囊藻属花朵但是此属的重要性在高纬度海洋碳循环。自然352:514 - 516 Sournia(1988)棕囊藻属(Prymnesiophyceae):但是此属物种多少?Nova Hedwigia 47:211 - 217 Treshnikov AF(1964)表面水循环在南冰洋。:俄罗斯。Sovet。Antarkticheskaia Eksped。,通知。biull。,45:5-8。Eng。transl。(1965):苏联南极探险、信息通报,5:81 - 83
Vaulot D, Birrien J-L玛丽·D Casotti R, Veldhuis MJW, aart Kraay GW Chretiennot-Dinet m j(1994)形态、倍性、颜料成分和培养菌株的基因组大小棕囊藻属(Prymnesiophyceae)。但是此属J Phycol 30:1022 - 1035
Veldhuis MJW Wassmann P (eds)(2005)开花动力学和生物防治的棕囊藻属,但是此属欧洲沿海水域的藻种的物种。有害藻类4:805 - 964
真实性PG,比利亚助教,Smayda TJ(1988)生态调查殖民棕囊藻属pouchetii但是此属的花朵。二世。生命周期终止现象盛开的角色。J浮游生物Res 10:749 - 766
公元前惠普尔SJ,彭定康,真实性PG(2005)生命周期的海洋藻类棕囊藻属,但是此属概念模型总结文献和指导研究。J 3月系统57:83 - 110 Zingone, Chretiennot-Dinet MJ,兰格M, Medlin路(1999)形态学和遗传特性的棕囊藻属cordata但是此属和p . jahnii (Prymnesiophyceae),两个新物种从地中海。J Phycol 39:1322 - 1337
生物地球化学(2007)83:19-27 DOI 10.1007 / s10533 - 007 - 9084 - 4
继续阅读:方法揭示的遗传多样性colonyforming prymnesiophytes棕囊藻属南极洲但是此属P globosa和P pouchetiipreliminary结果
这篇文章有用吗?